Лекция № 6: Изменения мысы соленого сырья. Созревание соленой продукции из животногосырья
Изменение массы соленого сырья
Наиболее существенным изменениям при посоле подвергается масса соленого сырья. Она может, как увеличиваться в процесса просаливания, преимущественно у животного сырья, так и уменьшаться.
Выход части воды в системе сырье : солевой раствор — диффузионно-осмотический процесс. При изучении закономерностей изменения массы при посоле у рыбы, которые с высокой степенью надежности можно распространить на все животное сырье в силу сходства химического состава и фракционного состава белков мышечной ткани, Леванидов И. П. отдавал предпочтение осмотической теории перемещения влаги. Ученый считал, что некоторые структурные элементы мышечной ткани рыб обладают свойствами полупроницаемых мембран, поэтому разность осмотических давлений в клетке и межклеточном пространстве является основной движущей силой влагопереноса.
Осмотическое давление разбавленных растворов описывается уравнением Вант-Гоффа:
 , (98)
, (98)
где  — изотонический коэффициент, составляет для хлорида натрия 2;
— изотонический коэффициент, составляет для хлорида натрия 2;
 — молярная концентрация растворенного вещества, моль/л;
— молярная концентрация растворенного вещества, моль/л;
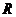 — универсальная газовая постоянная, Дж/(К·моль);
— универсальная газовая постоянная, Дж/(К·моль);
 — температура раствора, К.
— температура раствора, К.
Л.П. Миндер склонялся к тому, что влага из рыбы в основном извлекается за счет сил обычной молекулярной диффузии. Объясняя эту гипотезу тем, что мембраны у снулой рыбы из сплошных превращаются в сетчатые, которые проницаемы не только для воды, но и для органических веществ. Поэтому осмотические явления отсутствуют. Вероятно, оба процесса: осмос и диффузия имеют место при перемещении влаги из рыбы при ее просаливании.
Потеря влаги и поглощение соли мышечной тканью рыбы связаны обратно пропорциональной зависимостью. Таким образом, чем больше соли диффундировало в рыбу, тем меньше влаги в ней осталось. Следует отметить, что это справедливо при соленостях больше пяти процентов практически для всех видов животного сырья.
С другой стороны, в процессе хранения соленой рыбы в системе «рыба — тузлук» также имеют место массообменные процессы.
На I этапе хранения (до 25-30 суток в зависимости от солености рыбы, направляемой на посол) влага из рыбы удаляется. При последующем хранении (II этап) происходит впитывание влаги, так называемое набухание рыбы.
Процессы насыщения тканей рыбы водой в последующем хранении различные исследователи объясняют по-разному. Некоторые исследователи считают, что в результате гидролиза белков при посоле и последующем хранении происходит увеличение степени их дисперсности, что приводит к высвобождению связанной воды и ее диффузии из рыбы в тузлук (I этап). Размеры капилляров увеличиваются и уже не под действием осмотического давления, а за счет атмосферного давления происходит всасывание тузлука (II этап). Л.П. Миндер предполагает, что набухание вызвано двумя причинами: на первом этапе из рыбы успевает выйти больше воды, чем требуется для установления равновесия, следовательно, на втором этапе происходит поглощение тузлука (1 причина); пептизацией белков глобулинового характера в растворах низкой концентрации (2 причина). Отмечается также, что эти два процесса протекают одновременно и связаны между собой.
Представляет интерес следующее объяснение теории набухания. Перемещение воды в рыбу после завершения процесса влагоотдачи свидетельствует о нарушении электростатического равновесия в системе «рыба:тузлук». Это происходит за счет увеличения полярных групп при гидролизе белковых веществ и липидов и накопления заряженных аминных и карбоксильных групп и свободных аминокислот. Прекращение набухания при расщеплении белковых молекул в период созревания объясняется завершением процесса перераспределения слоев диполей воды около ионов Na+ и Cl–, причем, чем меньше расстояние между молекулами воды и ионами Na+ и Сl–, тем сильнее притяжение между ними. Набуханию более подвержено мясо нежирной рыбы, чем рыбы со значительным количеством жира в мышечной ткани.
Рассмотрим кинетику процессов извлечения воды из рыбы (I этап), которая характеризуется двумя периодами. В первом периоде происходит выделение межклеточного сока, который диффундирует через промежутки между волокнами. По мере выхода сока скорость выделения его в первый период уменьшается. Такое явление обычно происходит, когда концентрация соли в тканевом соке рыбы не превышает 12 %. После повышения концентрации соли во влаге рыбы выше 12 % солерастворимые белки тканей рыбы высаливаются, ткани сжимаются и уменьшаются в объеме. Вследствие этого увеличивается поперечное сечение промежутков между волокнами и уменьшается сопротивление выходу воды, влага выделяется более интенсивно, а скорость извлечения влаги уменьшается менее резко. Во второй период преимущественно извлекается внутриклеточный сок. Общее количество переместившейся воды из рыбы зависит от равновесной концентрации соли в ней.
На втором этапе больше влаги в рыбу в процессе ее хранения диффундирует при меньших концентрациях соли после посола. Это, видимо, можно объяснить тем, что при меньших концентрациях соли белки в меньшей степени высаливаются. Влага из клеток удаляется, в основном, за счет разности осмотического давления, при этом из рыбы удаляется больше воды, чем требуется для установления равновесия. На втором этапе хранения концентрация соли в клетке и тузлуке выравнивается за счет диффузии влаги в клетку опять-таки из-за разности осмотического давления. Эти процессы, по-видимому. протекают до полного выравнивания содержания соли как по всему объему рыбы, так и между рыбой и тузлуком. При увеличении концентрации соли во столько же раз увеличивается количество перемещающейся из рыбы воды. При постоянной концентрации соли в рыбе отношение количества перемещающейся воды к содержанию ее до просаливания является величиной постоянной, то есть чем больше начальное содержание воды в рыбе, тем больше ее выделяется при посоле. Скорость перемещения влаги из рыбы выше скорости перемещения соли в рыбу.
Если увеличивать температуру посола, то количество извлекаемой воды из рыбы при прочих равных условиях тоже увеличится.
Изменения массы рыбы при посоле обратимы не полностью. На эти изменения накладывают свой отпечаток начальные условия. Надо учитывать и изменения азотистых веществ и жира наряду с перемещениями воды и соли.
Потери плотных веществ при 0 °С больше, чем при 20 °С, но с увеличением плотности тузлука разница сглаживается. Это объясняется тем, что с повышением температуры усиливается высаливание белков, и потери плотных веществ меньше при теплом посоле, чем при холодном, а большую потерю массы при теплом посоле можно объяснить большей потерей воды.
Потери органических веществ в тузлуке при просаливании зависят и от продолжительности посола. При законченном посоле потери органических веществ выше, чем при прерванном посоле. Однако при дальнейшем хранении в тузлуке, потери органических веществ для рыбы законченного посола меньше, чем для рыбы прерванного посола.
Потери органических веществ у мороженой рыбы выше, чем у свежей. Такая закономерность особенно проявляется при низких температурах посола. По-видимому, это объясняется денатурационными изменениями белков, в результате чего нарушается связь белка и воды. Однако выход соленой продукции, приготовленной из мороженой рыбы, выше из3за меньших потерь воды.
Костные ткани в процессе посола также претерпевают изменения. Эти изменения зависят от минерализации костных тканей. Теоретически в случае абсолютно жесткой структуры кости при посоле должны увеличивать массу за счет насыщения солью воды, содержащейся в них. Однако при посоле тузлуком
(температура от 2 до 5 °С) у соленой трески масса костей увеличилась на 7,5%, а при посоле этой же трески в тузлуке с температурой от 7 до 13 °С масса костей уменьшилась на 1,5 %. Потери воды и плотных веществ, за исключением слабых посолов, больше, чем рыба приобретает соли. Поэтому масса соленой рыбы, как правило, меньше 100 %.
Тощие рыбы, как уже отмечалось, теряют в массе значительно больше, чем жирные. В свою очередь, чем крепче посол, тем больше рыба теряет воды и плотных веществ. Способ и техника посола также влияют на выход соленой рыбы.
Наибольший выход продукции дает смешанный посол, а наименьший — стоповый посол. При стоповом посоле нижние слои рыбы испытывают значительное давление, что ведет к уменьшению выхода рыбы. Влага, выделяющаяся из рыбы при стоповом посоле, свободно стекает, безвозвратно утрачиваясь. Также выход соленой рыбы уменьшается и при плотной ее укладке в таре.
Качество сырца также влияет на выход соленой рыбы. Рыба-сырец, посоленная в состоянии посмертного окоченения, теряет в массе меньше, чем рыба-сырец, находящаяся в стадии автолиза. При автолизе структура тканей рыхлая, соль при этом активно проникает к белкам и блокирует белковые молекулы, уменьшая их гидрофильность, вследствие этого большое количество влаги выделяется наружу.
Качество соли оказывает значительное влияние на выход соленой рыбы. Известно, что одновалентные металлы почти не вступают в связь с белками, поэтому Na+ не вступает во взаимодействие с белками рыбы. Двухвалентные ионы Са2+, Mg2+, которые могут встречаться в поваренной соли, наоборот, обладают высокой способностью вступать в соединение с белками, вызывая их денатурацию и уменьшая водоудерживающую способность. Если примесей в поваренной соли много, то выход продукции, обработанной такой солью, будет ниже.
Таким образом, на изменения массы и объема рыбы при посоле оказывают влияние следующие факторы: химический состав (жирность) рыбы, крепость тузлука и температура посола, способ и техника посола, механические воздействия, оказываемые на рыбу в процессе посола, качество сырья и качество используемой соли.
Дата добавления: 2018-04-15; просмотров: 426; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!