Адресная доставка полипептидов
Экзамен по биологии
I. Общая характеристика жизни
1)Биология, как комплексная наука: предмет и методы. Биология и медицина: точки соприкосновения. Здоровье и нездоровье в контексте иерархической структуры жизни.
Биология (от греч. bios — жизнь, logos — наука) — наука о жизни, об общих закономерностях существования и развития живых существ. Или иными словами, биологией называется наука, занимающаяся изучением жизни во всех ее проявлениях, а также свойств живого вообще. Предметом изучения биологии являются живые организмы, их строение, функции, развитие, взаимоотношения со средой и происхождение.
Термин «биология» ввел французский ученый эволюционист Жан Батист Ламарк в 1802 г.
Современная биология представляет собой систему наук о живой природе. Общие закономерности развития живой природы, раскрывающие сущность жизни, ее формы и развитие, рассматривает общая биология. Соответственно объектам изучения - животным, растениям, вирусам — существуют специальные науки, изучающие каждую из названных групп организмов.
По мере накопления фактического материала биология разделилась на несколько исследовательских областей. Биологические науки разделились по объектам исследования.
по систематическим категориям: вирусология, микробиология, бактериология ,ботаника, микология, зоология
по уровням организации живой материи: молекулярная биология, цитология, цитогенетика, морфология и физиология, экология, популяционная экология
|
|
|
в зависимости от изучаемых процессов: генетика, эмбриология, теория, этология, общая биология
Современная биология опирается на достижения других естественных наук — физики, химии, математики.
Основными частными методами в биологии являются:
• описательный;( Для того, чтобы выяснить сущность явлений, необходимо прежде всего собрать фактический материал и описать его)
• сравнительный;( позволяет путем сопоставления изучать сходство и различие организмов и их частей. На принципах этого метода была основана систематика и сделано одно из крупнейших обобщений - создана клеточная теория)
• исторический ; (выясняет закономерности появления и развития организмов, становления их структуры и функций.
• экспериментальный. ( связан с активным воздействием на них путем постановки опытов (экспериментов) в точно учитываемых условиях и путем изменения течения процессов в нужном исследователю направлении. Высшей формой эксперимента является моделирование изучаемых процессов.
| Уровень жизни | молекулярно-генетический | клеточный | организменный, онтогенетический | популяционно-видовой | биогеоценозный |
| элементарная структура | ген, сайт днк | клетка – элементарная единица жизни | особь (ее онтогенез) | популяция особей | биогеоценоз (экосистема) |
| элементарное явление | конвариантная репликация днк | Взаимодействующие потоки биоинформации, энергии и вещества | генотип ↓ фенотип | динамика гено(алле-ло)фондов, воспроиз-водство генотипов | вещественно – энергетические потоки и круговороты |
| вклад в здоровье, предпосылки нездоровья | сохранение биологически полезной биоинформации (здоровье). мутации – снижение качества биоинформации (нездоровье) | Вещественно-энергетическое и биоинформационное обеспечение жизнедеятельности(здоровье),дистрофии, малигнизация (нездоровье) | организм достаточной жизнеспособности (здоровье). ограниченная жизнеспособность, уродства, старение (нездоровье) | внутривидо-вые отношения – вклад в здоровье потомства. генетический груз (нездоровье) | межвидовые отношения и абиотические факторы здоровья в среде жизни. абиотические и биотические факторы нездоровья в среде жизни. |
Важность изучения биологии для медика определяется тем, что биология - это теоретическая основа медицины. Успехи медицины связаны с биологическими исследованиями, поэтому врач постоянно должен быть осведомлен о новейших достижениях биологии. Создание в XIX в. клеточной теории заложило подлинно научные основы связи биологии с медициной. В укреплении связей биологии с производством и медициной существенный вклад принадлежит генетике, данные которой имеют важнейшее значение в разработке основ диагностики, лечения и профилактики наследственных болезней. Открытие клетки и изучение микроскопического строения организмов позволили глубже понять причины возникновения болезненного процесса, способствовали разработке методов диагностики и лечения. В конце концов, и сам человек является живым организмом, поэтому биология является теоретической основой таких наук, как медицина, психология, социология и других.
|
|
|
|
|
|
2)Жизнь, как феномен материальности мира. Критика идеалистических и метафизических представлений о сущности жизни. Фундаментальные свойства жизни, как особого явления.
Жизнь - это макромолекулярная открытая система, для которой характерно определённая иерархическая организация, а также способность к воспроизведению, обмену веществ и тщательно регулируемому потоку энергии. Жизнь представляет собой распространяющийся центр упорядоченности в менее упорядоченной Вселенной.
|
|
|
Познание сущности жизни является одной из основных задач современной биологии. По мере развития науки постоянно происходило накопление фактического материала, который постепенно обобщался и осмысливался, что приводило к созданию различных теорий и гипотез. Однако не следует забывать, что любые философские обобщения и выводы всегда являются отражением мировоззрения конкретного ученного. Например, К. Линней внес много нового в биологию (например, предложил сохранившеюся поныне номенклатуру), но будучи глубоко религиозным человеком, стоял на позиции метафизики и признавал сотворение видов богом. По своему мировоззрению учёные с древнейших времен разделились на метафизиков и идеалистов. Материалисты признают, что весь мир материален, природа существует объективно – независимо от сознания человека, а сознание – продукт материи (мозга) и общественного развития. В противоположность к этому идеалисту утверждают, что первичным является нематериальное начало и что весь материальный мир - порождение сознания, духа. Связь идеализма с религией очевидна. Для биолога-идеалиста возникновение жизни на Земле – это вопрос о возникновении и источнике интеллекта – «мирового разума» и т.д.
Для материалистов, жизнь всецело материальна по своей природе; не она порождается интеллектом, а наоборот, интеллект возникает и является результатом прогрессивного развития материи.
Фундаментальные свойства живого:
• Обмен веществ
• Гомеостаз
• Рост и развитие
• Наследственность и изменчивость
• Дискретность и целостность
• Возбудимость и раздражимость
• Адаптивность
• Эволюционное развитие
• Онтогенез и филогенез.
3)Иерархические уровни организации жизни, обусловленные структурой эволюционного процесса. Элементарные единицы, элементарные явления и проявления главных свойств жизни на различных уровнях ее организации.
В основе лежит иерархический принцип, при котором существование жизни на каждом последующем уровне определяется и подготавливается структурой низшего уровня.
— Молекулярный и субклеточный уровень
— Клеточный уровень
— Тканевой и органный (органно-тканевой) уровень
— Организменный (онтогенетический) уровень
— Популяционно-видовой уровень
— Биоценотический и биосферный уровни
Фундаментальные свойства живого:
• Обмен веществ
• Гомеостаз
• Рост и развитие
• Наследственность и изменчивость
• Дискретность и целостность
• Возбудимость и раздражимость
• Адаптивность
• Эволюционное развитие
• Онтогенез и филогенез.
■ Молекулярный уровень
— основа живого — белки, состоящие всего из 20 аминокислот, и нуклеиновые кислоты, состоящие из 4 азотистых оснований
— у всех организмов биологическая энергия запасается в виде макроэргических фосфатов (АТФ, АДФ, АМФ)
— генетическая информация зашифрована в молекулярной структуре ДНК (у некоторых вирусов — в РНК)
— всем живым организмам на Земле присущ универсальный генетический код — каждой из двадцати аминокислот, образующих белки, соответствует определенная последовательность трех нуклеотидов в полинуклеотидной цепи
—элементарные структуры – центральные управляющие системы – коды наследственной информации, передаваемые от поколения к поколению - ген; элементарное явление – генные мутации
■ Клеточный уровень
— клетка является основной самостоятельно функционирующей элементарной биологической единицей, характерной для всех живых организмов
—элементарная структура – клетка; элементарное явление – деление клеток их развитие, клеточный метаболизм
■ Тканевой и органный (органно-тканевой) уровень
— совокупность клеток с одинаковым типом организации составляет ткань
— всего лишь 4 основных ткани входит в состав органов многоклеточных животных (эпителиальная, соединительная, мышечная, нервная)
— из тканей формируются органы, которым присущи определенные функции
—элементарная структура – ткань; элементарное эволюционное явление – дифференцировка и специализация клеток
—элементарная структура – орган; элементарное эволюционное явление – объединение различных тканей в общую структуру
■ Организменный (онтогенетический) уровень
— на данном уровне обнаруживается многообразие форм живых организмов
— на Земле обитает более миллиона видов животных организмов и около полумиллиона видов высших растений
— каждый вид состоит из отдельных индивидуумов — особей
— элементарная структура – особь, индивид; элементарное эволюционное явление – онтогенез
■ Популяционно-видовой уровень
— совокупность особей одного вида, населяющих одну территорию, свободно между собой скрещивающихся, составляет популяцию
— популяция — это элементарная единица эволюционного процесса — в ней начинаются процессы видообразования
— популяция входит в состав биогеоценозов
—элементарная структура – популяции любого вида, живых организмов; элементарное эволюционное явление – направленное изменение их генофонда
■ Биоценотический и биосферный уровни
— биогеоценозы — исторически сложившиеся устойчивые сообщества популяций различных видов, связанных между собой и с окружающей неживой природой обменом веществ, энергии и информации
— биогеоценозы составляют биосферу
—элементарная структура – биогеоценоз; элементарное эволюционное явление – вещественно-энергетический круговорот
— Биосферный.элементарная структура – биосфера; элементарное эволюционное явление – биогеохимический круговорот
II. Клеточный и молекулярно-генетический уровни организации жизни
1)Современное состояние клеточной теории, ее значение для биологии и медицины. Общие черты организации и отличительные особенности структурной организации про- и эукариотических клеток.
Клеточная теория сформулирована немецким исследователем, зоологом Т. Шванном (1839). Соавтором считают ботаника М. Шлейдена. Согласно теории, клетки являются структурной и функциональной основой живых существ. В конце 19 столетия немецкий патолог Р. Вирхов на основе новых фактов пересмотрел клеточную теорию. Ему принадлежит вывод о том, что клетка может возникнуть лишь из предшествующей клетки.
Современные положения клеточной теории:
o клетка является наименьшей биологической единицей живого, с помощью которой происходит извлечение из внешней среды, превращение и использование организмами энергии и веществ.
o клетки всех одноклеточных и многоклеточных организмов сходны по своему строению, химическому составу и важнейшим проявлениям жизнедеятельности.
o размножение клеток происходит путем деления исходной (материнской) клетки;
o клетки многоклеточных организмов специализированы: они выполняют разные функции и образуют ткани.
Со времён формулирования клеточной теории, ученными было сделано очень много открытий. И сейчас уже можно утверждать, что клетка является основной структурной и функциональной единицей организации живого.
Значение клеточной теории для медицины. Клетка - единица патологии. Клетку необходимо знать не только как единицу строения организма, но и как единицу патологических изменений. Практически все болезни связанные с нарушением структуры и функции клеток, из которых образуются все ткани и органы. Нарушение структуры и функции одних клеток является первопричиной возникновения и развития болезни, а нарушение других может быть уже следствием неблагоприятных изменений в организме. Например, при инфаркте миокарда нарушается функционирование, а потом наступает гибель кардиомиоцитов через острый недостаток кислорода. В силу того, что часть сердечной мышцы не берет участия в сокращении, нарушается кровоснабжение в организме, который приводит к гипоксии и изменениям функции и структуры клеток, в первую очередь нейронов головного мозга.
Прокариоты – доядерные организмы, не имеющие типичного ядра, заключенного в ядерную мембрану. К прокариотам относятся бактерии и синезеленые водоросли, объединённые общим понятием «дробянки».
Эукариоты – ядерные организмы, имеющие ядро, окруженное ядерной мембраной.
Независимо от индивидуальных структурно-функциональных особенностей все клетки одинаковым образом:
1. Хранят биологическую информацию
2. Редуплицируют генетический материал
3. Используют генетическую информацию для осуществления своих функций на основе синтеза белка
4. Хранят и переносят энергию
5. Превращают энергию (Е) в работу (А)
6. Регулируют обмен веществ.

2) Закономерности существования клетки во времени. Жизненный цикл клетки. Основное содержание и значение периодов жизненного цикла клетки
Каждая клетка должна осуществлять все процессы от которых зависит её жизнь, т.е. поглощать питательные вещества, использовать их как пластический и энергетический материал, избавляться от конечных продуктов жизнедеятельности и, наконец, воспроизводить самую себя. В результате этих процессов клетка постоянно изменяется, осуществляется её онтогенез, который называется жизненным циклом клетки, или клеточным циклом. Клеточный цикл — это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти.
Важным компонентом клеточного цикла является митотический (пролиферативный) цикл — комплекс взаимосвязанных и согласованных во времени событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления. Кроме того, в жизненный цикл включается период выполнения клеткой многоклеточного организма специфических функций, а также периоды покоя. В периоды покоя ближайшая судьба клетки не определена: она может либо начать подготовку к митозу, либо приступить к специализации в определенном функциональном направлении
Продолжительность митотического цикла для большинства клеток составляет от 10 до 50 ч. Длительность цикла регулируется путем изменения продолжительности всех его периодов. У млекопитающих время митоза составляет 1—1,5 ч, 02-периода интерфазы —2—5 ч, S-периода интерфазы — 6—10 ч.
Биологическое значение митотического цикла состоит в том, что он обеспечивает преемственность хромосом в ряду клеточных поколений, образование клеток, равноценных по объему и содержанию наследственной информации. Таким образом, цикл является всеобщим механизмом воспроизведения клеточной организации эукариотического типа в индивидуальном развитии.
Основные стадии митоза.
1. Редупликация (самоудвоение) генетической информации материнской клетки и равномерное распределение ее между дочерними клетками. Это сопровождается изменениями структуры и морфологии хромосом, в которых сосредоточено более 90 % информации эукариотической клетки.
2. Митотический цикл состоит из четырех последовательных периодов: пресинтетического (или постмитотического) G1, синтетического S, постсинтетического (или премитотического) G2 и собственно митоза. Они составляют автокаталитическую интерфазу (подготовительный период).
Фазы клеточного цикла:
1) пресинтетическая (G1). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки (гистоны, структурные белки, ферменты), РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов (т. е. структур, способных к ауторепродукции). Восстанавливаются черты организации интерфазной клетки после предшествующего деления;
2) синтетическая (S). Происходит удвоение генетического материала путем репликации ДНК. Она происходит полуконсервативным способом, когда двойная спираль молекулы ДНК расходится на две цепи и на каждой из них синтезируется комплементарная цепочка.
В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит из одной новой и старой цепи ДНК. Количество наследственного материала удваивается. Кроме этого, продолжается синтез РНК и белков. Также репликации подвергается небольшая часть митохонд-риальной ДНК (основная же ее часть реплицируется в G2 период);
3) постсинтетическая (G2). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных).
После этого наступает собственно митоз, который состоит из четырех фаз.
Деление клетки включает в себя два этапа – деление ядра (митоз, или кариокинез) и деление цитоплазмы (цитокинез).
Митоз состоит из четырех последовательных фаз – профазы, метафазы, анафазы и телофазы. Ему предшествует период, называемый интерфазой
Фазы митоза:
1) профаза. Центриоли клеточного центра делятся и расходятся к противоположным полюсам клетки. Из микротрубочек образуется веретено деления, которое соединяет центриоли разных полюсов. В начале профазы в клетке еще видны ядро и ядрышки, к концу этой фазы ядерная оболочка разделяется на отдельные фрагменты (происходит демонтаж ядерной мембраны), ядрышки распадаются. Начинается конденсация хромосом: они скручиваются, утолщаются, становятся видимыми в световой микроскоп. В цитоплазме уменьшается количество структур шероховатой ЭПС, резко сокращается число полисом;
2) метафаза. Заканчивается образование веретена деления.
Конденсированные хромосомы выстраиваются по экватору клетки, образуя метафазную пластинку. Микротрубочки веретена деления прикрепляются к центромерам, или кинетохо-рам (первичным перетяжкам), каждой хромосомы. После этого каждая хромосома продольно расщепляется на две хроматиды (дочерние хромосомы) которые оказываются связанными только в участке центромеры;
3) анафаза. Между дочерними хромосомами разрушается связь, и они начинают перемещаться к противоположным полюсам клетки со скоростью 0,2–5 мкм/мин. В конце анафазы на каждом полюсе оказывается по диплоидному набору хромосом. Хромосомы начинают деконденсироваться и раскручиваться, становятся тоньше и длиннее;
4) телофаза. Хромосомы полностью деспирализуются, восстанавливается структура ядрышек и интерфазного ядра, монтируется ядерная мембрана. Разрушается веретено деления. Происходит цитокинез (деление цитоплазмы). В животных клетках этот процесс начинается с образования в экваториальной плоскости перетяжки, которая все более углубляется и в конце концов полностью делит материнскую клетку на две дочерние.
При задержке цитокинеза образуются многоядерные клетки. Это наблюдается при размножении простейших путем шизогонии. У многоклеточных организмов так образуются синцитии – ткани, в которых отсутствуют границы между клетками (поперечно-полосатая мышечная ткань у человека).
Продолжительность каждой фазы зависит от типа ткани, физиологического состояния организма, воздействия внешних факторов (света, температуры, химических веществ) и пр.
3) Принцип структурно-функциональной организации генетического аппарата про- и эукариот. Понятие об уровнях структурно-функциональной организации эукариот – генном, хромосомном, геномном.
По химической организации материала наследственности и изменчивости эукариотические и прокариотические клетки принципиально не отличаются друг от друга. Генетический материал у них представлен ДНК. Общим для них является и принцип записи генетической информации, а также генетический код. Одни и те же аминокислоты шифруются у про- и эукариот одинаковыми кодонами. Принципиально одинаковым образом у названных типов клеток осуществляется и использование наследственной информации, хранящейся в ДНК. Однако некоторые особенности организации наследственного материала, отличающие эукариотические клетки от прокариотических, обусловливают различия в использовании их генетической информации.
Наследственный материал прокариотической клетки содержится главным образом в единственной кольцевой молекуле ДНК.
Наследственный материал эукариот больше по объему, чем у прокариот. Он расположен в основном в хромосомах, которые отделены от цитоплазмы ядерной оболочкой.
Значительные отличия имеются в молекулярной организации генов эукариотической клетки. В большинстве из них кодирующие последовательности экзоны прерываются интронными участками, которые не используются при синтезе тРНК, рРНК или пептидов. Эти участки удаляются из первично-транскрибируемой РНК, в связи с чем использование генетической информации в эукариотической клетке происходит несколько иначе. В прокариотической клетке, где наследственный материал и аппарат биосинтеза белка пространственно не разобщены, транскрипция и трансляция происходят почти одновременно. В эукариотической клетке эти два этапа не только пространственно отделены ядерной оболочкой, но и во времени их разделяют процессы созревания мРНК, из которой должны быть удалены неинформативные последовательности.
Строение гена у про- и эукариот.
| Прокариоты | Эукариоты |
| Ядро отсутствует и ДНК расположено свободно в цитоплазме | Ядро имеется и большая часть ДНК расположена в хромосомах. Небольшая часть ДНК расположена в цитоплазме |
| ДНК имеет слабое взаимодействие причем только с негистоновыми белками, обнаружены структурные единицы, близкие к нуклеосомам, но очень лабильные | ДНК расположена в ядре, взаимодействует с гистоновыми белками в 90% в постоянных нуклеосомах |
| ДНК двух цепочная, но кольцевая, называется генофор, содержит 106 пар нуклеотидов. | Ядра ДНК дуплекс спираль, у митохондрий дуплекс кольцо, напр., у человека 3*109 пар нуклеотидов. |
| Большую часть ДНК составляют последовательность соответствующая структурным генам. | |
| Репликация ДНК идет полуконсервативно, имеется один репликон. | Репликация ДНК тоже полуконервативная, но репликонов много |
| Единица транскрипции - оперон | Единица транскрипции – транскриптон |
| Одно звено метаболизма определяется группой структурных генов | Одно звено метаболизма определяется одним структурным геном. |
| Избыточного ДНК нет, 95% состоит из генов и только 5% - элементы дуплекса | Геном избыток у человека 56 % генов, кодируют участки, остальные не задействованыназываются - интроны |
Генный уровень организации наследственного материала. Ген – функционально неделимая единица. Участок молекулы ДНК. Совокупность генов данной клетки или особи в диплоидном наборе составляет генотип. Это работающая часть генов, которая выявляется по совокупности признаков в проявляющемся фенотипе. Гены относительно независимы друг от друга, поэтому возможны изменения отдельных признаков.
Хромосомный уровень организации наследственного материала. Связан с наличием у клеток эукариот особых надмолекулярных структур хромосом. Все гены в пределах одной хромосомы составляют одну группу сцепления. Этот уровень связан с перераспределением генов родителей у потомков при половом размножении.
Геномный уровень организации наследственного материала. Содержит гаплоидный объем наследственной информации достаточной для реализации морфофункциональной организации особи:
• обеспечивает определенный тип индивидуального развития,
• характеризуется видовой специфичностью,
• является результатом эволюции данного вида в конкретной среде.
Геномный уровень организации наследственного материала, объединяющий всю совокупность хромосомных генов, является эволюционно сложившейся структурой, характеризующейся относительно большей стабильностью, нежели генный и хромосомный уровни. На геномном уровне система сбалансированных по дозам и объединенных сложнейшими функциональными взаимосвязями генов представляет собой нечто большее, нежели простую совокупность отдельных единиц. Поэтому результатом функционирования генома является формирование фенотипа целостного организма. В связи с этим фенотип организма нельзя представлять как простую совокупность признаков и свойств, это организм во всем многообразии его характеристик на всем протяжении индивидуального развития. Таким образом, поддержание постоянства организации наследственного материала на геномном уровне имеет первостепенное значение для обеспечения нормального развития, организма и воспроизведения у особи в первую очередь видовых характеристик.
4) ДНК как генетический материал: соответствие структурно-химических характеристик биологическим функциям.
1. Днк – биополимер (макромолекулярный уровень);
2. Мономеры, строящие макромолекулу днк - нуклеотиды (4);
3. Нуклеотид состоит из азотистого основания (4), пятиуглеродного сахара дезоксирибозы и остатка фосфорной кислоты; разные нуклеотиды различаются азотистыми основаниями;
4. Азотистые основания: два – это пурины (аденин и гуанин), а два – это пиримидины (цитозин и тимин);
5. В макромолекуле днк нуклеотиды соединены химической связью между сахаром и фосфатом;
6. Биспираль днк (надмолекулярный уровень) взаимокомплементарные макромолекулы днк удерживаются водородными связями между пуринами и пиримидинами (а-т и г-ц); расстояния между основаниями в парах а-т и г-ц равны, что делает макромолекулы днк в биспирали антипараллельными; в паре а-т 2 водородных связи, в паре г-ц их 3.
7. Диаметр биспирали 2нм, расстояние в парах оснований 0,34нм, виток биспирали включает 10 пар нуклеотидов (в-спираль);
8. Наибольшую длину имеет биспираль днк хромосомы 1 человека (263 млн. П.н.), наименьшую – хромосомы 21 (50 млн. П.н.).
Структурно-химические характеристики, дающие возможность функционировать в качестве биоинформационного материала –
Функционально - генетическая задача структурно-химические характеристики
1. надежное сохранение биоинформации 1. химическая стабильность; биспираль-дублирование
биоинформации во взаимокомплементарных цепях;
исправление ошибок и репарация повреждений;
2. возможность записать большой объем 2. биополимер, в структуре которого допускается любая
биоинформации последовательность мономеров (4 нуклеотида);
3. передача биоинформации в ряду клеточных 3. надмолекулярная структура в виде биспирали из взаимо-
поколений без существенных потерь комплементарных цепей (матричный синтез, репликация);
4. использование биоинформации для 4. то же (матричный синтез, транскрипция);
организации клеточных функций;
5. регуляция генетических функций днк 5. у эукариот – нуклеогистоновый комплекс
с возможностью химических взаимодействий гистонов с
кислыми негистоновыми белками (транскрипционные
факторы), “спирализация- деспирализация” биспирали,
химическая модификация молекул днк (метилирование);
6. определенный объем «информационного 6. ошибки репликации, рекомбинации, репарации дей
Шума» в виде генных (истинных) мутаций, ствие мутагенов.
дающих новую биоинформацию.
5) Репликация ДНК как матричный процесс: инициация, элонгация, терминация. Репликация ДНК прокариот и митохондрий. Репликация концевых участков (теломеры) молекулы ДНК.
репликация – способность к самокопированию
Репликация ДНК - это построение двух новых (дочерних) молекул ДНК на основе одной старой (родительской) молекулы с помощью матричного синтеза по принципу комплементарности. Новые молекулы состоят из одной старой и одной новосинтезированной нитей ДНК и почти ничем не отличаются от старой родительской молекулы, т.е. являются её точными копиями ("репликами").
Как матричный процесс репликация включает 3 фазы:
1. инициация или начало
2. элонгация или наращивание (удлиннение) строящейся дочерней макромолекулы (цепи) днк
3. терминация или завершение
Инициация. Репликация ДНК начинается в специфических местах называемых точками начала репликации (origin). Чем больше размер генома организма, тем большее количество точек репликации ему необходимо. Происходит в результате взаимодействия ориджин репликации со специфическими белками. Они переходят в такое состояние, при котором они способны связывать фермент ДНК-полимеразу В И.р. участвуют также другие белки: ДНК-геликаза (расплетающий фермент: расплетает ДНК на две отдельные цепи). Образуется репликативная вилка. На разъединение комплементарной связи в каждой паре нуклеотидов тратится энергия 2-х молекул АТФ. В одной репликативной вилке действуют две геликазы, которые движутся в противоположных направлениях. У эукариот одновременно образуется тысячи таких вилок, что обеспечивает высокую скорость репликации, топоизомераза (ДНК-топоизомеразы, ДНК-релаксирующие ферменты: суперскрученность убирается), белок, связывающийся с одноцепочечной ДНК (SSB).
Элонгация.
Элонгация – рост цепи иРНК (Скорость примерно 50 нуклеотидов/секунду), происходит неодинаково для двух ее цепей. ДНК- полимераза III прокариот и δ- или α-ДНК-полимеразы эукариот осуществляют синтез лишь в направлении 5’>3’. Цепь с такой направленностью – лидирующая ДНК-полимераза III или δ-полимераза непрерывно присоединяют к ней комплементарные нуклеотиды. Цепь с полярностью 3’>5’ является отстающей и достраивается по частям (также в направлении 5’>3’). α-ДНК-полимераза (или ДНК-полимераза III) синтезирует на этом цепи короткие участки - фрагменты Оказаки. Синтез фрагментов Оказаки и лидирующей цепи начинается с образования РНК-праймеров (затравок) длиной 10-15 рибонуклеотидов ферментом праймазой (РНК-полимеразой). Ни одна из ДНК- полимераз не способна начать синтез ДНК с нуля, а может лишь достраивать существующую цепь. Параллельно с образованием лидирующей цепи или фрагментов Оказаки происходит удаление рибонуклеотидов из праймеров и замена их нуклеотидами ДНК (при участием β-ДНК-полимеразы, которая имеет как экзонуклеазную, так и полимеразную активность). Все ДНК-овые фрагменты сшиваются лигазой.
Терминация.
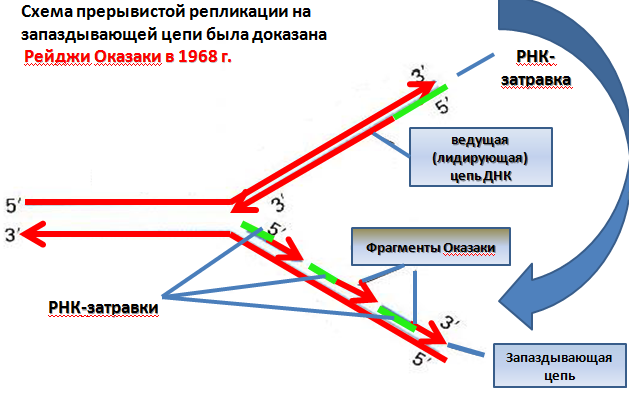 Терминация (завершение) репликации происходит тогда, когда пробелы между фрагментами Оказаки заполнятся нуклеотидами (при участии ДНК-лигазы) с образованием двух непрерывных двойных цепей ДНК и когда встретятся две репликативные вилки. Затем происходит закручивание синтезированных ДНК с образованием суперспиралей.
Терминация (завершение) репликации происходит тогда, когда пробелы между фрагментами Оказаки заполнятся нуклеотидами (при участии ДНК-лигазы) с образованием двух непрерывных двойных цепей ДНК и когда встретятся две репликативные вилки. Затем происходит закручивание синтезированных ДНК с образованием суперспиралей.

Репликация днк прокариотических клеток: особенности
1. Репликация происходит с одной точки инициации одним блоком или репликоном, не прерываясь, с образованием двух “репликационных вилок”;
2. Ключевой фермент репликации днк у прокариот – днк-полимераза iii, функционирующая в комплексе с примерно 20 белками;
3. Днк-полимераза iii катализирует синтез как лидирующей, так и отстающей полинуклеотидных цепей днк;
4. На завершающей стадии заполнение брешей на месте разрушенных праймеров (запаздывающая цепь) происходит с участием фермента днк-полимеразы i;
5.терминация репликации у прокариот происходит, когда “репликационная вилка” достигает участка днк с особыми сайтами ter
6. Днк-полимераза ii участвует в процессах молекулярной репарации повреждений прокариотической днк;
Репликация митохондриальной днк: особенности
1. Репликация митохондриальной днк не связана с периодом s интерфазы клеточного цикла и происходит в любой его временной точке, за исключением непосредственно митоза;
2. Мднк отдельных органелл реплицируется независимо от репликации днк других митохондрий клетки; за клеточный цикл днк одних органелл реплицируется более одного раза, а других – не реплицируется вовсе;
3. Репликация митохондриальной днк катализируется ферментом γ(гамма)днк-полимеразой;
4. Для каждой из макромолекул (полинуклеотидных цепей) биспирали митохондриальной днк есть своя точка инициации;
5. Образование рнк-праймеров при репликации митохондриальной днк катализируется ферментом днк-зависимой рнк-полимеразой.
Репликация теломерных (концевых) участков молекул днк эукариотических клеток: особенности
1. Когда “репликативная вилка” достигает конца линейной молекулы эукариотической ДНК, на которой как на матрице происходит репликация отстающей дочерней цепи днк, не остается места для образования рнк-праймера, с которого мог бы начаться синтез днк терминального фрагмента оказаки, вследствие чего с каждым циклом репликации молекулы днк укорачиваются (маргинотомия днк) – у человека на 50-100 нуклеотидов;
2. Укорочения удается избежать благодаря двум моментам: - теломерная ДНК образована нуклеотидными повторами, богатыми гуаниловым нуклеотидом (человек - gggtta); - специальные нуклеотидные последовательности “узнаются” ферментным теломеразным комплексом со свойствами обратной транскриптазы, который “достраивает” соответствующие концевые фрагменты днк на своей рнк - матрице; Теломераза — фермент, добавляющий особые повторяющиеся последовательности ДНК (TTAGGG у позвоночных) к 3'-концу цепи ДНК на участках теломер, которые располагаются на концах хромосом в эукариотических клетках.
3. Теломеразный комплекс активен в интенсивно размножающихся клетках
6) ДНК как генетический материал. Репарация генетического материала, виды репарации.
1. Днк – биополимер (макромолекулярный уровень);
2. Мономеры, строящие макромолекулу днк - нуклеотиды (4);
3. Нуклеотид состоит из азотистого основания (4), пятиуглеродного сахара дезоксирибозы и остатка фосфорной кислоты; разные нуклеотиды различаются азотистыми основаниями;
4. Азотистые основания: два – это пурины (аденин и гуанин), а два – это пиримидины (цитозин и тимин);
5. В макромолекуле днк нуклеотиды соединены химической связью между сахаром и фосфатом;
6. Биспираль днк (надмолекулярный уровень) взаимокомплементарные макромолекулы днк удерживаются водородными связями между пуринами и пиримидинами (а-т и г-ц); расстояния между основаниями в парах а-т и г-ц равны, что делает макромолекулы днк в биспирали антипараллельными; в паре а-т 2 водородных связи, в паре г-ц их 3.
7. Диаметр биспирали 2нм, расстояние в парах оснований 0,34нм, виток биспирали включает 10 пар нуклеотидов (в-спираль);
8. Наибольшую длину имеет биспираль днк хромосомы 1 человека (263 млн. П.н.), наименьшую – хромосомы 21 (50 млн. П.н.).
Репарация генетических повреждений – свойство живых организмов восстанавливать нарушения и повреждения, возникшие в ДНК в результате ошибок репликации, а также при воздействии разнообразных эндогенных и внешних мутагенных факторов. Одной из причин возникновения целого ряда наследственных заболеваний и раковых опухолей является нарушение репарации ДНК.
Источники повреждения ДНК:
· Ультрафиолетовое излучение
· Радиация
· Химические вещества
· Ошибки репликации ДНК
· Апуринизация — отщепление азотистых оснований от сахарофосфатного остова
· Дезаминирование — отщепление аминогруппы от азотистого основания
Каждая из систем репарации включает следующие компоненты:
§ ДНК-геликаза — фермент, «узнающий» химически изменённые участки в цепи и осуществляющий разрыв Н-связей вблизи от повреждения;
§ фермент, удаляющий повреждённый участок;
§ ДНК-полимераза — фермент, синтезирующий соответствующий участок цепи ДНК взамен удалённого;
§ ДНК-лигаза — фермент, замыкающий последнюю связь в полимерной цепи и тем самым восстанавливающий её непрерывность.
Разновидности:
1. Эксцизионная или дорепликативная (с вырезками)
В настоящее время известно два типа эксцизионной репарации:
ü Эксцизия азотистых оснований с помощью специальных ферментов – гликозилаз с последующим восстановлением нативной структуры ДНК;
ü Эксцизия нуклеотидов из цепи ДНК. После удаления поврежденных нуклеотидов из цепи ДНК происходит ее застройка помощью ДНК-полимеразы I.
Эксцизионная репарация представляет собой многоэтапный процесс и включает:
1)«Узнавание» тиминового димера.
2)Инцизию - надрезание одной цепи ДНК вблизи димера.
3)Эксцизию - удаление сегмента ДНК с поврежденными нуклеотидами (тиминовым димером).
4)Ресинтез ДНК.
5)Восстановление непрерывности репарируемой цепи за счет образования фосфодиэфирных связей.
2. Пострепликативная (путем рекомбинации)
Пострепликативная репарация происходит тогда, когда в ДНК возникает так много повреждений, что в ходе эксцизионной репарации клетка не успевает их полностью устранить, а также, если повреждены гены, контролирующие синтез ферментов, участвующих в эксцизионной репарации. В результате после репликации такой ДНК в дочерней цепи на месте повреждений, имеющихся в материнской нити, образуются «бреши». Этот способ восстановления целостности ДНК заключается в репарации пробелов, образующихся в дочерних цепях напротив не удаленных в ходе репликации димеров. Основная часть таких пробелов репарируется путем рекомбинационных обменов между двумя сестринскими цепями. В процессе используются ферменты ДНК- полимераза I и лигаза, и белок RecA.
3. Световая репарация (самопроизвольное устранение нарушений под действием видимого света).
7) Ген как функциональная единица наследственности. Свойства гена. Классификация генов. Особенности организации генов у про- и эукариот. Генетический код как способ записи наследственной информации, его свойства. Цистрон, его структура.
Ген – функционально неделимая единица, определяет строение белковой полипептидной цепи
участок молекулы ДНК, кодирующий развитие отдельного признака.. Совокупность генов данной клетки или особи в диплоидном наборе составляет генотип. определяющий строение белковой полипептидной цепи
Свойства генов:
1. дискретность — несмешиваемость генов;
2. стабильность — способность сохранять структуру;
3. лабильность — способность многократно мутировать;
4. множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
5. аллельность — в генотипе диплоидных организмов только две формы гена;
6. специфичность — каждый ген кодирует свой признак;
7. плейотропия — множественный эффект гена;
8. экспрессивность — степень выраженности гена в признаке;
9. пенетрантность — частота проявления гена в фенотипе;
10. амплификация — увеличение количества копий гена.
Классификация:
1. Структурные гены — уникальные компоненты генома, представляющие единственную последовательность, кодирующую определённый белок или некоторые виды РНК.
2. Функциональные гены — регулируют работу структурных генов. Промотор, оператор, энхансер, сайленсер, терминатор

прокариоты
Оперон – это группа следующих подряд структурных генов, находящихся под контролем одного регуляторного участка ДНК. Все сцепленые гены оперона кодируют ферменты одного метаболического пути (например, расщепление лактозы).
Организация по типу оперона позволяет бактериям быстро переключать метаболизм с одного субстрата на другой. Бактерии не синтезируют ферменты определенного метаболического пути в отсутствии необходимого субстрата, но способны начать их синтезировать при появлении субстрата.
Структура генов эукариот
Благодаря экзонно – интронной организации генов создаются предпосылки для альтернативного сплайсинга. Альтернативний сплайсинг - процесс «вырезания» разных интронов из первичного РНК-транскрипта в результате чего на основе одного гена могут синтезироватся разные белки. Явление альтернативного сплайсинга имеет место у млекопитающих при синтезе различних антител на основе иммуноглобулиновых генов.
Генетический код - это способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
В ДНК используется четыре азотистых основания. В РНК используются те же нуклеотиды (аденин, гуанин, цитозин), за исключением тимина, который заменён похожим нуклеотидом — урацилом (У). В молекулах ДНК и РНК нуклеотиды выстраиваются в цепочки и, таким образом, получаются последовательности генетических букв.
Белки практически всех живых организмов построены из аминокислот всего 20 видов. Эти аминокислоты называют каноническими. Каждый белок представляет собой цепочку или несколько цепочек аминокислот, соединённых в строго определённой последовательности. Эта последовательность определяет строение белка, а, следовательно, все его биологические свойства.
Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов (кодоны-терминаторы в ген.коде иРНК: УАА, УГА, УАГ). Набор из трёх нуклеотидов называется триплетом.
Свойства генетического кода:
1. Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). При этом 4 нуклеотида (А,Т,Г,Ц) образуют 64 кодона, из которых 61 кодирует 20 аминокислот.
2. Непрерывность — триплеты считываются без пропусков.
3. Неперекрываемость — один нуклеотид входит в состав только одного триплета.
4. Однозначность (специфичность) — один кодон кодирует только одну аминокислоту.
5. Вырожденность (избыточность) — одну аминокислоту может кодировать несколько триплетов (от 2 до 6, исключение Метионин и триптофан – 1 триплет).
6. Универсальность — у всех живых существ одни триплеты кодируют одинаковые аминокислоты.
Ген как функциональную единицу предложено называть цистроном. Именно цистрон определяет последовательность аминокислот в каждом специфическом белке. Цистрон, в свою очередь, подразделяется на предельно малые в линейном измерении единицы - реконы, способные к рекомбинации при кроссинговере. Выделяют, кроме того, мутоны - наименьшие части гена, способные к изменению (мутированию). Размеры рекона и мутона могут равняться одной или нескольким парам нуклеотидов, цистрона - сотням и тысячам нуклеотидов. Цистрон – это последовательность нуклеотидов ДНК, которая определяет отдельную генетическую функцию полипептидной цепи. Ген может быть представлен одним или несколькими цистронами. Сложные гены содержащие в себе несколько цистронов называются полицистронными.
8) Уровни структурно-функциональной организации эукариот. Генный уровень, его характеристика.
Принципы структурно-функциональной организации генетического аппарата прокариот и эукариот различны. В генетическом аппарате эукариотических клеток выделяют три уровня структурно-функциональной организации – генный, хромосомный и геномный.
На каждом из уровней решаются свои специфические задачи, чем обеспечиваются известные варианты наследования признаков, формы генотипической и фенотипической изменчивости. На этом уровне изучается структура молекулы ДНК, биосинтез белка и др.
На генном уровне решаются специфические функционально-генетические задачи –
Наследственность:
1. Моногенное, независимое наследование признаков;
2. Независимое комбинирование отдельных признаков в фенотипе потомства;
Передача генов от родителей потомку необходима для развития у него определенных признаков
Изменчивость:
Независимое изменение структуры (нуклеотидной последовательности днк) отдельных генов – генные или истинные мутации – мутационная генотипическая изменчивость;
Функционально-генетической единицей генного уровня является ген
Ген - участок днк, кодирующий аминокислотную последовательность в белке-ферменте. В геноме h.s. На долю структурных генов приходится не более 5% днк. Функции порядка 50% днк неизвестны – эгоистичная днк;
“нуклеотидная последовательность” = “ген”, но без точной характеристики функции, “цистрон” = “ген”, применяется в генетике прокариот.
Структура эукариотического гена
единицей транскрипции у эукариот является транскриптон, функция которого определяется наличием следующих элементов:
§ неинформативная зона: промотор + инициатор
§ информативная зона: структурный ген, разделенный на экзоны и интроны
§ терминатором
Работу транскриптона регулируют несколько генов-регуляторов, дающих или кодирующих синтез нескольких белков-репрессоров. Потому что индукторами эукариот являются сложные молекулы для расщепления которых требуется несколько ферментов.
Промотор – особый участок в молекуле ДНК, который указывает место начала транскрипции.
Оператор – определенные нуклеотидные последовательности ДНК, предшествующие структурной части регулируемого гена. Включает и выключает работу структурных генов.
Терминатор – участок, где прекращается дальнейший рост цепи РНК и происходит ее освобождение от матрицы ДНК.
Репрессор – регуляторный белок, подавляющий транскрипцию генов регулируемого им оперона в результате связывания с оператором (регуляторным участком оперона).
Индуктор – небольшая эффекторная молекула, связывающаяся с регуляторным белком, или физический фактор (свет, температура), которые стимулируют экспрессию генов, находящихся в неактивном состоянии.
Благодаря наличию генного уровня возможно индивидуальное, раздельное (дискретное) и независимое наследование и изменения отдельных признаков.
9) Структура и виды РНК, роль РНК в процессе реализации наследственной информации.
Молекула РНК полимер мономерами которой являются нуклеотиды. РНК представляет собой однонитевую молекулу, состоящую из азотистого основания, пентозы и фосфорной кислоты. Три азотистых основания такие же как у ДНК, но вместо тимина - урацил. Содержание РНК в клетках сильно колеблется. Оно заметно повышено в клетках в которых происходит синтез белка.
Виды РНК:
1) Транспортная РНК (т-РНК). Молекулы т – РНК самые короткие: они состоят всего из 80-100 нуклеотидов. Транспортная РНК в основном содержится в цитоплазме клетки. Функция состоит в переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетки на долю т-РНК приходится 10%.
2) Рибосомная РНК (р-РНК). Это самые крупные РНК: в их молекулу входит 3-5 тысяч нуклеотидов. Р-РНК составляет существенную часть структуры рибосомы. Из общего содержания РНК клетки на долю р-РНК приходится 90%
3) Информационная РНК или матричная. Содержится в ядре и цитоплазме. Функция её состоит в переносе информации о структуре белка в рибосомах. На долю и-РНК приходится примерно 0,5-1% от общего содержания РНК клетки.
Матричная, или информационная, РНК (мРНК, или иРНК). Транскрипция. Для того чтобы синтезировать белки с заданными свойствами, к месту их построения поступает «инструкция» о порядке включения аминокислот в пептидную цепь. Эта инструкция заключена в нуклеотидной последовательности матричных, или информационных РНК (мРНК, иРНК), синтезируемых на соответствующих участках ДНК. Процесс синтеза мРНК называют транскрипцией.
Образуемая в ходе транскрипции мРНК содержит точную копию информации, записанной в соответствующем участке ДНК. Тройки рядом стоящих нуклеотидов мРНК, шифрующие аминокислоты, называют кодонами. Последовательность кодонов мРНК шифрует последовательность аминокислот в пептидной цепи. Кодонам мРНК соответствуют определенные аминокислоты.
Транспортная РНК (тРНК). Трансляция. Важная роль в процессе использования наследственной информации клеткой принадлежит транспортной РНК (тРНК). Доставляя необходимые аминокислоты к месту сборки пептидных цепей, тРНК выполняет функцию трансляционного посредника.
Молекулы тРНК представляют собой полинуклеотидные цепи, синтезируемые на определенных последовательностях ДНК. Они состоят из относительно небольшого числа нуклеотидов —75—95. В результате комплементарного соединения оснований, которые находятся в разных участках полинуклеотидной цепи тРНК, она приобретает структуру, напоминающую по форме лист клевера.
В ней выделяют четыре главные части, выполняющие различные функции. Акцепторный «стебель» образуется двумя комплементарно соединенными концевыми частями тРНК. Он состоит из семи пар оснований. 3'-конец этого стебля несколько длиннее и формирует одноцепочечный участок, который заканчивается последовательностью ЦЦА со свободной ОН-группой. К этому концу присоединяется транспортируемая аминокислота. Остальные три ветви представляют собой комплементарно спаренные последовательности нуклеотидов, которые заканчиваются неспаренными участками, образующими петли. Средняя из этих ветвей — антикодоновая — состоит из пяти пар нуклеотидов и содержит в центре своей петли антикодон. Антикодон — это три нуклеотида, комплементарные кодону мРНК, который шифрует аминокислоту, транспортируемую данной тРНК к месту синтеза пептида.
Первичная структура тРНК, определяемая последовательностью нуклеотидов, формирует вторичную структуру тРНК, имеющую форму листа клевера.
Наследственная информация, «записанная» в молекулах ДНК и «переписанная» на мРНК, расшифровывается в ходе трансляции благодаря двум процессам специфического узнавания молекулярных поверхностей. Сначала фермент аминоацил-тРНК-синтетаза обеспечивает соединение тРНК с транспортируемой ею аминокислотой. Затем аминоацил-тРНК комплементарно спаривается с мРНК благодаря взаимодействию антикодона с кодоном. С помощью системы тРНК язык нуклеотидной цепи мРНК. транслируется в язык аминокислотной последовательности пептида.
Рибосомная РНК (рРНК). Рибосомный цикл синтеза белка. Процесс взаимодействия мРНК и тРНК, обеспечивающий трансляцию информации с языка нуклеотидов на язык аминокислот, осуществляется на рибосомах. Последние представляют собой сложные комплексы рРНК и разнообразных белков, в которых первые образуют каркас. Рибосомные РНК являются не только структурным компонентом рибосом, но и обеспечивают связывание их с определенной нуклеотидной последовательностью мРНК. Этим устанавливаются начало и рамка считывания при образовании пептидной цепи. Кроме того, они обеспечивают взаимодействие рибосомы и тРНК. Многочисленные белки, входящие в состав рибосом наряду с рРНК, выполняют как структурную, так и ферментативную роль.
Рибосомы про- и эукариот очень сходны по структуре и функциям. Они состоят из двух субчастиц: большой и малой. У эукариот малая субчастица образована одной молекулой рРНК и 33 молекулами разных белков. Большая субчастица объединяет три молекулы рРНК и около 40 белков. Прокариотические рибосомы и рибосомы митохондрий и пластид содержат меньше компонентов.
В рибосомах имеется две бороздки. Одна из них удерживает растущую полипептидную цепь, другая — мРНК. Кроме того, в рибосомах выделяют два участка, связывающих тРНК. В аминоацильном, А-участке размещается аминоацил-тРНК, несущая определенную аминокислоту. В пептидильном, П-участке располагается обычно тРНК, которая нагружена цепочкой аминокислот, соединенных пептидными связями. Образование А- и П-участков обеспечивается обеими субчастицами рибосомы.
В каждый момент рибосома экранирует сегмент мРНК протяженностью около 30 нуклеотидов. При этом обеспечивается взаимодействие только двух тРНК с двумя расположенными рядом кодонами мРНК .Трансляция информации на «язык» аминокислот выражается в постепенном наращивании пептидной цепи в соответствии с инструкцией, заключенной в мРНК. Этот процесс протекает на рибосомах, которые обеспечивают последовательность расшифровки информации с помощью тРНК. В ходе трансляции можно выделить три фазы: инициацию, элонгацию и терминацию синтеза пептидной цепи.
Фаза инициации, или начало синтеза пептида, заключается в объединении двух находящихся до этого порознь в цитоплазме субчастиц рибосомы на определенном участке мРНК и присоединении к ней первой аминоацил-тРНК. Этим задается также рамка считывания информации, заключенной в мРНК.
Фаза элонгации, или удлинения пептида, включает в себя все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Она представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил-тРНК очередного кодона, находящегося в А-участке, комплементарное взаимодействие между антикодоном и кодоном.
Фаза терминации, или завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ или У ГА), когда тот входит в зону А-участка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы
10) Транскрипция биоинформации с ДНК на РНК как матричный процесс – фазы инициации, элонгации, терминации. Регуляция генной активности у про- и эукариот.
Транскрипция – это матричный процесс образования в соответствии с принципом комплементарности молекул РНК на матричной полинуклеотидной цепи биспирали ДНК;
Биоинформационно транскрипция является процессом “переписывания” или “переноса” части ДНК-текста на РНК с целью организации в цитоплазме синтеза полипептида с соответствующей аминокислотной последовательностью. Т.о. – это промежуточное звено, необходимое для перевода ДНК-биотекстов (генотип) в белковые биотексты (фенотип);
В эукариотических клетках три РНК-полимеразы. РНК- полимераза i – синтез пре-р-РНК, РНК-полимераза ii – синтез пре-и(м)РНК, транскрипция структурных (смысловых полипептидных) генов с последующей трансляцией, РНК-полимераза iii - транскрипция генов тРНК.
У эукариот транскрипция днк происходит частями, т.е. Неодновременно: - по времени (онтогенез), - по месту (тип клеток), - по интенсивности.
В силу экзон/интронной структуры генов эукариот сначала образуется первичный продукт – пре-и(м)РНК транскрипт, затем процессинг.
Фаза инициации
У эукариот в области промотора транскриптона, включающего “знаковые” (узнаваемые белками) нуклеотидные последовательности ( - тата -; - цаат -), образуется мультибелковый инициаторный комплекс. Среди белков комплекса – общие транскрипционные факторы, обусловливающие точную “посадку” на биспираль ДНК РНК-полимеразы, определение точки начала транскрипции и положения инициирующего кодона, взаимодействие с энхансерами и специфическими транскрипционными факторами. В месте “посадки” РНК-полимеразы биспираль ДНК расплетается и ее матричная цепь открывается для считывания биоинформации;
Осуществляется:
· ДНК – зависимыми РНК – полимеразами
· Оператором
· Промотором, содержащим блок Прибнова 5' - ТАТААТ - 3', который является стартом транскрипции
· Белковыми факторами инициации
Фаза элонгации
Наращивание РНК транскрипта идет в направлении 5́ → 3́; скорость транскрипции – 30 нуклеотидов/сек. “надежность” транскрипции – 1 ошибка на 2·104 включенных нуклеотида;
РНК-полимераза наращивает молекулу РНК транскрипта на 3́ конце путем присоединения из пула рибонуклеозидтрифосфатов – АТФ, ГТФ, УТФ и ЦТФ – требуемого рибонуклеозидмонофосфата;
У прокариот еще до окончания транскрипции на РНК транскрипте с 5́ конца начинается трансляция. У эукариот первоначально образуется пре-и(м)РНК транскрипт, который отсоединяется от матрицы и подвергается процессингу с образованием зрелых и(м)РНК. пре-РНК транскрипты образуются для рРНК и, возможно, для тРНК.
Регуляторы скорости транскрипции: энхансеры – (ускоряют) и сайленсеры (замедляют)
Фаза терминации
Процесс транскрипции завершается, когда РНК-полимераза, перемещаясь вдоль матричной цепи ДНК, достигает кодона-терминатора (АТТ, АТЦ, АЦТ).
По завершении транскрипции эукариотические и(м)РНК на 3́ конце полиаденилируются;
По завершении транскрипции пре-и(м)РНК транскрипт (пре-рРНК транскрипт и, возможно, пре-тРНК транскрипт) отсоединяется от матрицы ДНК и подвергается процессингу.
Терминация транскрипции осуществляется палиндромом, который образует шпилечную структуру или фигуру «креста»
Регуляция генной активности:
Эукариоты
Различают неспецифическую и специфическую регуляцию генной активности;
Механизм неспецифической регуляции работает на всех уровнях организации генетического аппарата (генном, хромосомном, геномном) и заключается в гетерохроматизации генетического материала;
Главные участники специфической регуляции генной активности: промотор, энхансеры и сайленсеры, общие и специфические транскрипционные факторы, белки ядерного матрикса, метилирование азотистых оснований днк (цитозина);
“ген-господин” контролирует синтез ключевого белка-регулятора экспрессии многих генов (“гены-рабы”).
Регуляция генной активности идет за счет специальных регуляторных белков, которые кодируются регуляторными генами
Прокариоты
Для прокариот характерен полицистронный (цистрон = структурный ген) принцип организации генной активности. Цистроны объединяются в единую функциональную систему- оперон. Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований если они участвуют в одной цепи биохимических преобразований.
Лактозный оперон e. Coli. Негативная регуляция - в отсутствие в среде лактозы ген-оператор (часть промотора) заблокирован белком-репрессором (продукт гена-регулятора). Появившаяся лактоза инактивирует репрессор. Ген-оператор активируется, РНК-полимераза “допускается” к промотору и оперон транскрибируется; позитивная регуляция – основана на наличии белка-активатора (cap), который увеличивает сродство промотора к РНК-полимеразе и активирует оперон, но в условиях высокой концентрации цамф (ситуация глюкозного “голода”, необходим переход к использованию лактозы и некоторых др. Сахаров).
11) Транкриптон: единица генетической (транскрипционной) активности ДНК у эукариот.
Единица генетической активности ДНК у эукариот – транскриптон

Инициирующий транскрипцию кодон (ТАЦ –Метионин), за которым следует первый значимый Кодон;
Экзоны (функциональная часть: транскрибируются, транслируются)
Интроны ( НЕинформативные участки: транскрибируются, НЕтранслируются)
Завершающий транскрипцию кодон-терминатор (АТТ, АТЦ или АЦТ), следующий за последним значимым кодоном;
12) Посттранскрипционные процессы – процессинг пре-и(м)РНК (гяРНК) транскрипта, сплайсинг гяРНК, ядерно-цитоплазматический транспорт и(м)РНК, ядерные и цитоплазматические информосомы.
После окончания транскрипции, как у прокариот, так и у эукариот происходит цепь биохимических реакций, которая приводит к созреванию молекул предшественников: транспортной РНК(пре-тРНК) и рибосомной РНК (пре-рРНК) и пре-мРНК (только у эукариот). Совокупность реакций, приводящая к формированию зрелой (готовой к трансляции) молекулы мРНК, называется процессингом.
Особенности транскрипции у эукариот. Процессинг мРНК. Процессннг включает следующие преобразования молекулы мРНК:
1) метилирование и кэпирование;
2) полиаденилирование;
3) сплайсинг.
Эукариотические мРНК несут, как правило, на 5'-конце дополнительную группу: КЭП-модифицированный в 7-положении метилированный остаток гуанозин-5'-трифосфата, соединенный с концевым нуклеозидом 5'-5'-способом, Кэпирование РНК осуществляется ферментами: гуанилтрансферазой и метилтрансферазой. Предполагают, что КЭП необходим для регуляции трансляции и для стабилизации мРНК, (он предохраняет ее от действия 5'-экзонуклеаз), К 3'-концу РНК после завершения ее синтеза с помощью фермента поли(А)-полимеразы присоединяется последовательность полиадениловой кислоты. Этот процесс называют полиаденилированием. Остальные варианты преобразования пре-мРНК: вырезание нитронов и сшивание экзонов (сплайсинг) в эукариотических генах, а также образование различных сочетаний экзонов, входящих в зрелую мРНК (альтернативный сплайсинг) .
Эукариотические мРНК в отличие от прокариотических стабильны в течение часов и суток. Это объясняется, во-первых, стабилизацией 5'- и 3'-концов, а во-вторых, связыванием мРНК с белками (т.е. образованием информосом). Пре-мРНК на всех стадиях процессинга и после него связана с белками. Информосомы могут быть ядерными и цитоплазматическими. Ядерные информосомы - это рибонуклеопротеиновые (РНП) частицы с константой седиментации 30S.
Посттранскрипционный внутриядерный перенос пре-мРНК из ядра в цитоплазму осуществляется с помощью ядерных информосом. При этом переносе зрелой мРНК происходит замена связанных с мРН К белков. Ядерные информоферы (белковые глобулы) остаются в ядре, а мРНК после перехода в цитоплазму объединяется с новыми белками, образуя цитоплазматические информосомы. Цитоплазматические информосомы не обязательно транслируются, т.е. могут быть свободными.
Пре-и(м)рнк транскрипт эукариот содержит транслируемую (соответствует экзонам ДНК) и нетранслируемую (соответствует интронам ДНК) части;
В области сплайосом транскрипт разрывается и интронные участки удаляются;
Экзонные участки пре-и(м)РНК транскрипта соединяются – сплайсинг – с образованием зрелых и(м)РНК;
Зрелые и(м)РНК комплексируются с белками (информоферы) – ядерные информосомы, которые перемещаются к ядерной оболочке (возможно, к порам);
Сплайсинг - вырезание интронов и сшивание экзонов (сплайсосомы –мярнк + белки) идет в ядре
Альтернативный сплайсинг - путем различных комбинаций экзонных участков из одного пре-и(м)РНК транскрипта образуется несколько зрелых и(м)РНК.
Кепирование – на 5,-конце мрнк образуется защита – кэп(шапочка) путем + метилированного гуанозина ч/з остаток фосфорной кислоты)
Полиаденилирование – на 3, конце мрнк защита – «поли (а)хвост» идет + адениловых остатков (до200)
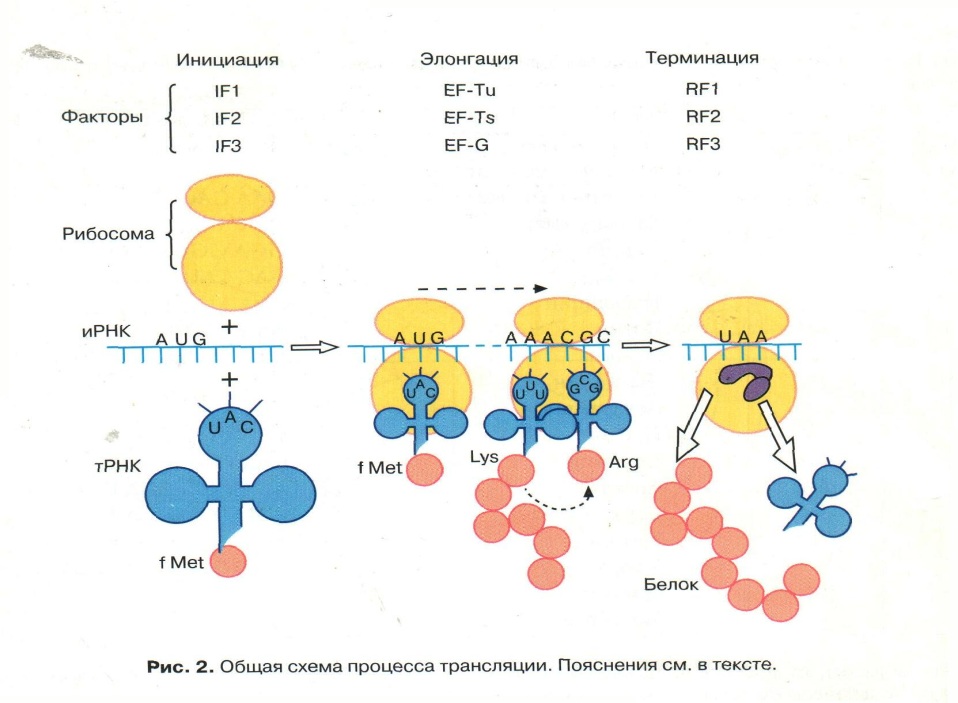
13) Трансляция биоинформации – рибосомный цикл. Биосинтез белка.
Трансляция - перевод нуклеотидной последовательности мРНК в аминокислотную последовательность полипептида. Осуществляется в цитоплазме на рибосомах (на шЭПС, в гиалоплазме) Участвуют: мРНК, рРНК, тРНК, белковые факторы
В названном комплексе и(м)РНК несет биоинформацию о последовательности аминокислот в полипептиде; рибосомы создают условия для пространственного взаиморасположения участников процесса и для осуществления требуемых каталитических и регуляторных актов; тРНК “транслирует” нуклеотидный текст в аминокислотный;
Трансляция, начавшись, идет, не прерываясь, вплоть до завершения; субъединицы рибосом, тРНК, некоторые белки участвуют в процессе неоднократно – “рибосомный цикл биосинтеза белков”; начало и окончание трансляции задаются однозначно сигнальными кодонами и(м)РНК – инициирующим и терминатором;
Трансляция – матричный процесс; в нем выделяют фазы инициации, элонгации и терминации;
Фаза инициации
1. Включение в пептидную цепь в качестве первой а. К. – модифицированного МЕТИОНИНА, с которого начинается ВСЕ синтезы пептидов.
2. Инициаторная тРНК с а.к. - метионин узнает малую субъединицу рибосом с помощью Белковых Факторов Инициации
3. Малая субъединица рибосом узнает мРНК и ее инициирующий кодон стартовый - АУГ
4. Образуется комплекс Малая субъединица рибосом + тРНК + мРНК
5 . Формируются участки ФЦР: А – центр, где устанавливается связь «кодон –антикодон» и П –центр, где образуются пептидные связи между аминокислотами.
Фаза элонгации
Фаза элонгации – циклический процесс: с каждым очередным шагом строящийся полипептид удлинняется на один аминокислотный остаток;
Влючает в себя 3 этапа.
На первом этапе молекула аминоацил-тРНК связывается со свободным участком рибосомы, примыкающим к занятому Р-участку. Связывание осуществляется путем спаривания трех нуклеотидов антикодона с тремя нуклеотидами иРНК, находящимися в А-участке.
На втором этапе карбоксильный конец полипептидной цепи отделяется в Р-участке от молекулы тРНК и образует пептидную связь с аминокислотой, присоединенной к молекуле тРНК в А-участке.Соединение аминокислот в цепи возможно потому, что у каждой из них имеются две разные химические группы: обладающая основными свойствами аминогруппа, NH2, и кислотная карбоксильная группа, СООН. Карбоксильная группа одной аминокислоты может образовать амидную (пептидную) связь с аминогруппой другой аминокислоты.
На третьем этапе новая пептидил-тРНК переносится в Р-участок рибосомы, в то время как рибосома продвигается вдоль молекулы иРНК ровно на три нуклеотида. Таким образом, в результате элементарного элонгационного цикла полипептид удлиняется на одну аминокислоту.
События повторяются до момента, когда в а-центре рибосомы появится кодон-терминатор и(м)рнк (уаа, уаг, уга), для которого нет трнк с комплементарным антикодоном;
Фаза терминации
Завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ или У ГА), когда тот входит в зону А-участка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы.
Прокариотическая клетка
Принципиально процесс трансляции у прокариот следует той же схеме, что и у эукариот;
Рибосомы прокариот имеют меньшие размеры – 70s (80s), также как малая – 30s (40s) и большая -50s (60s) субъединицы; в рибосомальных субъединицах прокариот меньше белков – 21 (30) в малой и 34 (45) в большой;
Хотя у прокариот цистороны одного оперона транскрибируются единым болоком, процесс трансляции происходит поцистронно: каждый цистрон имеет отдельный инициирующий кодон ( триплет аминокислоты формилметионина) и кодон-терминатор; хотя трансляция у прокариот происходит поцистронно, синтез разных полипептидов оперона регулируется единым образом;
14) Посттрансляционные процессы – приобретение пептидами третичной и четвертичной структур. Адресный транспорт полипептидов, детекция и уничтожение функционально дефектных полипептидов. Регуляция количества образуемых белков.
Последовательность аминокислотных остатков в полипептиде составляет его первичную (линейную,одномерную) структуру, определяемую последовательностью нуклеотидов в гене непосредственно;.
Функциональная активность белков в клетке и/или организме связана с приобретением третичной (трехмерной, объемной) структуры путем фолдинга (сворачивания), а также с объединением простых белков (полипептидов) в ди-/мультимерные гомо- или гетеробелковые комплексы.
Предположительно фолдинг может происходить
· случайно путем образования объемной структуры до момента, когда находится вариант с минимальной свободной энергией; расчеты говорят, что на это необходимы млн. Лет, в клетке это занимает минуты;
· Модульный принцип требует участия специальных белков – либо ферментов,либо молекулярных шаперонов(Молекулярные шапероны – семейство специализированных белков, обеспечивающих быстрое нахождение полипептидом правильной трехмерной структуры (фолдинг) и адресную доставку полипептидов в органеллу, например, в митохондрию;
Шапероны участвуют в фолдинге, не определяя трехмерную структуру; это функция аминокислотной последовательности;
Адресная доставка полипептидов
Полипептиды (простые белки) образуются на полисомах в цитоплазме; полисомы - свободные и прикрепленные к мембранам цитоплазматической сети; структура и функции митохондрий обеспечиваются полипептидами, образуемыми на полисомах цитоплазмы под контролем ядерных генов, и в самой органелле под контролем собственных генов;
Полипептиды, транспорт которых связан с канальцами цитоплазматической сети, имеют лидерную последовательность (25±11 аминокислот). Она вместе с полипептидом после идентификации цитоплазматическими “узнающими сигнал” частицами прикрепляется к мембранному рецептору цитоплазматической сети с образованием поры – транслокона. Попав через пору в просвет, полипептид гидролизуется с присоединением олигосахаридов и в транспортных пузырьках переносится в комплекс гольджи;
В комплексе гольджи полипептиды “сортируются” на секретируемые и предназначенные для внутриклеточных структур; первые – разделяются на “порции” или “кванты”, “упаковываются” в оболочку и в пузырьках доставляются к плазмалемме; вторые – метятся в зависимости от органеллы;
Белки, предназначаемые для лизосом, узнаются в комплексе гольджи с образованием метки. Такие помеченные полипептиды фиксируются на рецепторах мембраны комплекса с последующим образованием пузырьков, несущих исключительно лизосомальные ферменты.
Перемещение митохондриальных белков, образующихся под контролем ядерных генов на свободных полисомах цитоплазмы, требует их комплексирования с шаперонами. В комплексе с последними они проникают через 2 мембраны в матрикс органеллы; здесь при участии других шаперонов эти белки приобретают трехмерную конфигурацию. Белки узнаются органеллой благодаря наличию во внешней мембране рецептора, а в полипептиде лидерной аминокислотной последовательности; если полипептид предназначается для межмембранного пространства митохондрии, он имеет 2 лидерных последовательности: одна – транспорт в матрикс, вторая – транспорт через внутреннюю мембрану;
Дата добавления: 2019-09-13; просмотров: 269; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!