СПОРОГЕНЕЗ И ГАМЕТОГЕНЕЗ У РАСТЕНИЙ
Особенности гаметогенеза у растений. Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор; 2-й этап — гаметогенез — включает образование зрелых гамет.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) —мега- или макроспорогенезом.
Микроспорогенез протекает аналогично сперматогенезу у животных, вплоть до образования сперматид, а мегаспорогенез соответствует образованию ооцита П.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У растений отсутствует зачатковый путь в том смысле, как это имеет место у животных.
Микроспорогенез и микрогаметогенез. Рассмотрим микроспорогенез и микрогаметогенез на наиболее общем примере покрытосеменных растений. В субэпидермальной ткани молодого пыльника, называемой археспорием, каждая клетка после ряда делений становится материнской клеткой пыльцы, которая проходит все фазы мейоза.
52
В результате двух мейотических делений возникают четыре гаплоидные микроспоры (рис. 24, /). Они лежат четверками, их называют тетрадами спор.
При созревании тетрады распадаются на отдельные микроспоры. Этим заканчивается микроспорогенез.
Вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
|
|
|
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, не способны к движению и называются спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три клетки. Две из них — спермин и одна вегетативная.
Мегаспорогенез и мегагаметогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, часто только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры (см. рис._ 24, 2).
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады гаплоидна. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития). Судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
|
|
|
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти, и затем ее ядро претерпевает ряд митозов. При этом сама клетка не делится, она образует зародышевый мешок.
У разных растений число митозов может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) бывает три деления, в результате возникает восемь одинаковых ядер. Во время этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других в противоположном конце зародышевого мешка, называемого халазальным (рис. 25). В дальнейшем ядра обособляются в самостоятельные клетки, имеющие значительное количество цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка и две так называемые синергиды — обра-
53

 зуют яйцевой аппарат. Синергиды играют вспомогательную роль при оплодотворении, они скоро разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное, или центральное, ядро зародышевого мешка. Оставшиеся у халазального конца зародышевого мешка три ядра обособляются в клетки; они называются антиподами. Антиподы, так же как и синергиды, играют вспомогательную роль при развитии зиготы и вскоре разрушаются.
зуют яйцевой аппарат. Синергиды играют вспомогательную роль при оплодотворении, они скоро разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное, или центральное, ядро зародышевого мешка. Оставшиеся у халазального конца зародышевого мешка три ядра обособляются в клетки; они называются антиподами. Антиподы, так же как и синергиды, играют вспомогательную роль при развитии зиготы и вскоре разрушаются.
|
|
|
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 одинаковых гаплоидных ядер, из которых только одно образует яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно.
Кроме моноспорического типа развития, который только что рассмотрен, существуют и другие. При различных типах развития зародышевого мешка сохраняется разное'1 количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям (если сохраняются две мегаспоры — биспорический; четыре — тетраспорический тип развития).
Сравнение процессов созревания половых клеток у животных и растений (рис. 23 и 24) показывает почти полный параллелизм их, несмотря на то, что расхождение (дивергенция) растений и животных в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов в растительном и животном мире.
|
|
|
ОПЛОДОТВОРЕНИЕ
Сущность оплодотворения. Оплодотворением принято называть побуждение яйца к развитию путем объединения в нем ядер (кариогамии) мужских и женских половых клеток. Оплодотворение представляет собой необратимый процесс; однажды оплодотворенное яйцо не может быть оплодотворено вновь. Син-гамия (слияние мужских и женских половых клеток) и кариогамия составляют сущность процесса оплодотворения.
Оплодотворение у животных. Процесс оплодотворения у животных можно разделить на несколько фаз. Первая фаза начинается с того, что сперматозоид либо прикрепляется к любой точке поверхности яйцеклетки, либо проникает в нее через микропиле. Момент соприкосновения головки сперматозоида с яйцом является начальным в цепи химических реакций. Эту фазу называют фазой активации яйца.
Вторая фаза процесса оплодотворения начинается после проникновения в яйцо одного (моноспермия), а у некоторых животных и нескольких сперматозоидов (полиспермия). Проникший сперматозоид «готовится» к слиянию с женским ядром и последующему митозу: ядро сперматозоида постепенно набухает и приобретает вид интерфазного ядра. Такое ядро называют семенным или мужским пронуклеусом. Ядро яйцеклетки, прошедшей все фазы мейоза, готовое к слиянию с ядром сперматозоида, называют женским пронуклеусом.
К моменту соприкосновения сперматозоида с яйцом и проникновения его внутрь ядро яйца у разных животных может находиться на разных стадиях деления созревания. Кариогамия же может произойти только после окончания мейоза, т. е. после образования женского пронуклеуса.
В акте оплодотворения два гаплоидных пронуклеуса сливаются в одно ядро, образуя зиготу. Этот момент является кульминационным пунктом процесса полового размножения. В результате кариогамии гомологичные хромосомы, разошедшиеся в мейозе у предыдущего поколения, вновь воссоединяются в одном ядре зиготы. Так восстанавливается диплоидный набор хромосом при половом размножении.
В цитоплазму яйцеклетки у млекопитающих проникает не только головка (ядро) сперматозоида, как это считали раньше, но и его шейка и даже хвостовая часть, что создает возможность передачи некоторого количества цитоплазмы мужского организма потомству.
Оплодотворение у растений. Оплодотворение у растений в принципе сходно с таковым у животных, но имеет и некоторые особенности.
Раньше было сказано, что микрогаметогенез завершается образованием двух спермиев, которые формируются или в пыльцевом зерне, или в пыльцевой трубке при прорастании пыльцевого зерна.
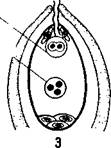
Пыльцевая трубка, дорастая до микропиле зародышевого мешка, соприкасается с яйцевым аппаратом — яйцеклеткой и синергидами (рис. 26, /). При соприкосновении конца пыльцевой трубки с синергидами пыльцевая трубка лопается, а синергиды разрушаются. Передвигающиеся по пыльцевой трубке по мере ее роста два генеративных ядра — спермин после разрыва трубки вместе с ее содержимым попадают внутрь зародышевого мешка (рис. 26, 2). Из двух проникших в зародышевой мешок спермиев один спермин сливается с гаплоидным ядром яйцеклетки (рис. 26, 3). Слияние ядра спермия с ядром яйцеклетки является собственно оплодотворением у растений. В оплодотворенной яйцеклетке — зиготе восстанавливается диплоидное число хромосом. Из зиготы развивается зародыш семени.
54
55

|
|
|
|
|

| Лыльиебая трудка ^ Синергиды Яйцеклетка |
| ВегетатиЬное ядро Спрр ' Мии |
| Зигота Триплоидное ядро шдоспврма |
| ' ентральнш ядра |
| Anmunodi |
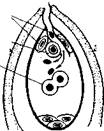
| 26. Схема двойного оплодотворения у растений: 1 — проникновение пыльцевой трубки в зародышевый мешок; 2 — излияние содержимого пыльцевой трубки в зародышевый мешок; 3 — зародышевый мешок после оплодотворения. |
Следует отметить, что у растений, так же как и у животных, женские ядра перед проникновением спермиев в зародышевый мешок могут находиться на разных стадиях, а кариогамия происходит лишь после завершения мейоза.
У покрытосеменных растений, кроме зародыша, в семени развивается дополнительный эмбриональный орган — эндосперм, который представляет собой питательное депо зародыша. Начало развития эндосперма обеспечивается вторым оплодотворением. Второй спермий пыльцевой трубки, попадая в зародышевый мешок, сливается с диплоидным ядром центральной клетки зародышевого мешка. При этом образуется клетка с тройным набором хромосом: два одинаковых набора хромосом материнского организма и один набор отцовского (рис. 26, 3). Слияние одного спермия с яйцеклеткой, а другого с ядром-центральной клетки называют двойным оплодотворением. Это открытие сделано в 1898 г. нашим соотечественником С. Г. Навашиным.
Таков в самых общих чертах процесс оплодотворения у животных и растений. Однако он подвержен приспособительным изменениям в зависимости от особенностей строения половых клеток и биологии размножения, свойственных каждому виду животных и растений.
До сих пор рассматривался процесс оплодотворения, связанный с поведением ядер, но, очевидно, этим суть оплодотворения не исчерпывается: оно представляет собой сложный физиологический и биохимический процесс.
Моноспермия, полиспермия, избирательность и селективное оплодотворение. Несмотря на огромное количество сперматозоидов и пыльцевых зерен, приходящихся на одну яйцеклетку животного и растения, оплодотворение осуществляется, как правило, лишь при участии одного сперматозоида и одного пыльцевого зерна. Такой тип оплодотворения называют моноспермным
56
оплодотворением. Он характерен для большинства животных и растений.
Моноспермное оплодотворение контролируется рядом механизмов. Одним из них является блокирование яйца после проникновения в него одного сперматозоида. Блокирование яйцеклеток у некоторых животных протекает в течение минуты и обеспечивается образованием оболочки оплодотворения и возникновением перивителлинового пространства. Физиологическая сущность блокирования яйца после соприкосновения головки сперматозоида с поверхностью яйцеклетки не выяснена.
У растений также после проникновения в зародышевый мешок одной пыльцевой трубки развиваются процессы, препятствующие проникновению других.
Однако у ряда животных в цитоплазму яйцеклетки проникает несколько сперматозоидов. Явление проникновения в цитоплазму яйцеклетки нескольких сперматозоидов называется полиспермией. Полиспермия широко распространена у беспозвоночных: моллюсков, иглокожих, насекомых; встречается она и у позвоночных: рыб (закономерно у акуловых), амфибий, рептилий и птиц. У млекопитающих в норме полиспермия встречается редко (1—2%); исключением являются однопроходные, у которых это явление, возможно, является правилом. У растений также наблюдаются случаи полиспермии, когда в зародышевый мешок проникает несколько пыльцевых трубок. Полиспермия отмечена у свеклы, хлопчатника, гречихи, табака и других растений.
Однако, несмотря на проникновение в яйцеклетку нескольких сперматозоидов в случае полиспермии, с женским пронук-леусом соединяется только один мужской пронуклеус. Остальные элиминируются. Полиспермная кариогамия, т. е. слияние нескольких пронуклеусов, в норме не обнаружена. Описаны случаи, когда у растений дополнительные спермин сливаются не с ядром яйцеклетки, а с другими ядрами зародышевого мешка (синергидами или антиподами), тогда из одного зародышевого мешка развивается несколько зародышей (полиэмбриония).
Проникновение в цитоплазму яйцеклетки нескольких спермиев и слияние с ее ядром только одного из них давно наводили на мысль, что данный процесс не является чисто механическим. Допускается возможность избирательности в процессе кариогамии, т. е. способность женского пронуклеуса сливаться с определенным мужским пронуклеусом.
Яйцеклетки могут преимущественно оплодотворяться сперматозоидами с определенными свойствами так же за счет конкуренции последних. Такое явление называется селективным оплодотворением. Оно приводит к ограничению свободного скрещивания (панмиксии) и является одним из приспособительных механизмов изоляции в эволюции растений и животных.
57
Дата добавления: 2020-01-07; просмотров: 2443; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!