Неорганические вещества клетки
| Вода |
Вода — самое распространенное в живых организмах неорганическое соединение. Ее содержание колеблется в широких пределах: в клетках эмали зубов вода составляет по массе около 10%, а в клетках развивающегося зародыша — более 90%.
Без воды жизнь невозможна. Она не только обязательный компонент живых клеток, но и среда обитания организмов. Биологическое значение воды основано на ее химических и физических свойствах.
| Физические и химические свойства воды |
Химические и физические свойства воды необычны. Они объясняются, прежде всего, малыми размерами молекул воды, их полярностью и способностью соединяться друг с другом водородными связями.
В молекуле воды один атом кислорода ковалентно связан с двумя атомами водорода. Молекула полярна: кислородный атом несет частичный отрицательный заряд, а два водородных — частично положительные заряды. Это делает молекулу воды диполем. Поэтому при взаимодействии молекул воды друг с другом между ними устанавливаются водородные связи. Они слабее ковалентной, но, поскольку каждая молекула воды способна образовывать 4 водородные связи, они существенно влияют на физические свойства воды. Большая теплоемкость, теплота плавления и теплота парообразования объясняются тем, что большая часть поглощаемого водой тепла расходуется на разрыв водородных связей между ее молекулами. Вода обладает высокой теплопроводностью. Вода практически не сжимается, прозрачна в видимом участке спектра. Наконец, вода — единственное вещество, плотность которого в жидком состоянии больше, чем в твердом.
| Биологическое значение воды |
Физические и химические свойства делают ее уникальной жидкостью и определяют ее биологическое значение.
© Вода — хороший растворитель ионных (полярных) соединений, а также некоторых не ионных, в молекуле которых присутствуют заряженные (полярные) группы. Если энергия притяжения молекул воды к молекулам какого-либо вещества больше, чем энергия притяжения между молекулами вещества, то молекулы гидратируются и вещество растворяется (рис. 256). По отношению к воде различают:
¨ гидрофильные вещества — вещества, хорошо растворимые в воде;
¨ гидрофобные вещества — вещества, практически нерастворимые в воде.
|
 Рис 254. Свойства молекулы воды:
1 — когезия молекул воды; 2 — гидратация катиона; 3 — гидратация аниона.
Рис 254. Свойства молекулы воды:
1 — когезия молекул воды; 2 — гидратация катиона; 3 — гидратация аниона.
© Большая теплоемкость и теплопроводность воды препятствуют возникновению "горячих точек" в организме, так как способствуют равномерному распределению тепла в клетке.
© Благодаря большой теплоте испарения воды, происходит охлаждение организма.
© Плотность льда меньше плотности воды. Поэтому при замерзании водоемов подо льдом остается жизненное пространство для водных организмов.
© Благодаря силам адгезии[7] и когезии[8], вода обладает свойством капиллярности, то есть способности подниматься по капиллярам (один из факторов, обеспечивающих движение воды в сосудах растений) (рис. 254).
© Вода является непосредственным участником многих химических реакций (гиролитическое расщепление белков, углеводов, жиров и др.).
© Несжимаемость воды определяет напряженное состояние клеточных стенок (тургор), а также выполняет опорную функцию (гидростатический скелет, например, у круглых червей).
| Минеральные вещества |
Минеральные вещества клетки в основном представлены солями, которые диссоциируют на анионы и катионы, некоторые — в неионизированной форме в микродозах (Fe, Mg, Cu, Co, Ni и др.)
Для процессов жизнедеятельности клетки наиболее важны катионы Na+, Ca2+, Mg2+, анионы HPO42-, Cl-, HCO3-. Концентрации ионов в клетке и среде ее обитания, как правило, различны. Например, во внешней среде (плазме крови, морской воде) K+ всегда меньше, а Na+ всегда больше, чем в клетке. Существует ряд механизмов, позволяющих клетке поддерживать определенное соотношение ионов в протопласте и внешней среде.
Различные ионы принимают участие во многих процессах жизнедеятельности клетки:
© катионы К+, Na+, Ca2+ обеспечивают раздражимость живых организмов;
© катионы Mg2+, Mn2+, Zn2+, Ca2+ и др. необходимы для нормального функционирования многих ферментов;
© образование углеводов в процессе фотосинтеза невозможно без Mg2+ (составная часть хлорофилла);
© слабощелочная реакция содержимого клетки поддерживается анионами слабых кислот (НСО3-, НРО4-) и слабыми кислотами (Н2СО3);
© От концентрации солей внутри клетки зависят ее буферные свойства. Буферностью называют способность клетки поддерживать слабощелочную реакцию своего содержимого на постоянном уровне. Внутри клетки буферность обеспечивается главным образом анионами H2PO4- и НРО42-. Во внеклеточной жидкости и в крови роль буфера играют Н2СО3- и НСО32-.
Фосфатная буферная система:
Низкий pH Высокий pH
НРО42- + Н+ ←―――――――→H2PO4-
Гидрофосфат — ион Дигидрофосфат — ион
Бикарбонатная буферная система:
Низкий pH Высокий pH
НСО3- + Н+ ←―――――――→H2СO3
Гидрокарбонат — ион Угольная кислота
Некоторые неорганические вещества содержатся в клетке не только в растворенном, но и в твердом состоянии. Например, Са и Р содержатся в костной ткани, в раковинах моллюсков в виде двойных углекислых и фосфорнокислых солей.
Органические вещества
Органические соединения составляют в среднем 20–30 % массы клетки живого организма. К ним относятся биологические полимеры — белки, нуклеиновые кислоты и полисахариды, а также жиры и ряд низкомолекулярных органических веществ — аминокислоты, простые сахара, нуклеотиды и т.д. Различные типы клеток содержат разное количество органических соединений. Так, растительные клетки богаты углеводами, а животные — белками (40–50 % в животной, 20–35 % в растительной). Каждая из групп органических веществ в клетках любого типа выполняет сходные функции.
Белки
Из органических веществ клетки по количеству и значению на первом месте стоят белки. Белки, или протеины (от греч. протос — первый, главный), — высокомолекулярные органические вещества, характеризующиеся строго определенным элементарным составом и распадающиеся при гидролизе до аминокислот.
В состав белков входят (в %): углерод — 50-55, водород -6,5-7,3, азот — 15-18, кислород — 21-24, сера — до 2,4 и зола — до 5,5. Часть белков образует комплексы с другими молекулами, содержащими фосфор, железо, цинк и медь.
Белки обладают большой молекулярной массой: молекулярная масса альбумина (одного из белков яйца) — 36000, гемоглобина — 152000, миозина (одного из белков мышц) — 500000. Один из белков — глобулин молока — имеет молекулярную массу 42000. Его формула С1864Н3012О576N468S21. Существуют белки, молекулярная масса которых в 10 и даже в 100 раз больше. Для сравнения: молекулярная масса спирта — 46, уксусной кислоты — 60, бензола — 78.
| Аминокислотный состав белков |
Белки являются полимерами, то есть состоят из нескольких структурных единиц — мономеров. Белки представляют собой непериодичные полимеры, мономерами которых являются аминокислоты.
В клетках и тканях обнаружено свыше 170 различных аминокислот, но в состав белков входит лишь 26. Причем 6 из них являются нестандартными. Они образуются в результате модификации стандартных аминокислот уже после их включения в полипептидную цепь. Поэтому обычными компонентами белков можно считать лишь 20 аминокислот.
В зависимости от того, могут ли аминокислоты синтезироваться в организме, различают:
© заменимые аминокислоты — десять аминокислот, синтезируемых в организме;
© незаменимые аминокислоты — аминокислоты, которые в организме не синтезируются.
Растения способны синтезировать все необходимые им аминокислоты, а животные же — лишь половину. Незаменимые аминокислоты должны поступать в организм вместе с пищей.
В зависимости от аминокислотного состава, белки бывают:
© полноценными, если содержат весь набор аминокислот;
© неполноценными, если какие-то аминокислоты в их составе отсутствуют.
Основная масса аминокислот, входящих в состав молекулы белка, яв-
|
 Рис. 255. Общая формула аминокислот.
Рис. 255. Общая формула аминокислот.
В зависимости от количества аминогрупп и карбоксильных групп, входящих в состав аминокислот, различают:
© нейтральные аминокислоты, имеющие одну карбоксильную группу и одну аминогруппу;
© основные аминокислоты, имеющие более одной аминогруппы;
© кислые аминокислоты, имеющие более одной карбоксильной группы.
Свойства аминокислот зависят не столько от количества аминогрупп и карбоксильных групп, сколько от их радикалов. Радикалы могут быть простыми и сложными, небольшими по размерам и громоздкими, гидрофильными и гидрофобными, химически инертными и высокоактивными, полярными и неполярными, заряженными положительно и отрицательно и т.д. Особенности радикалов, их число и расположение оказывают существенное влияние на структуру, химические и физические свойства полипептидов.
Аминокислоты являются амфотерными соединениями, так как в растворе они могут выступать как в роли кислот, так и оснований. В водных растворах аминокислоты существуют в разных ионных формах. Это зависит от рН раствора и от того, какая аминокислота: нейтральная, кислая или основная.
| Пептиды |
Пептиды — органические вещества, состоящие из остатков аминокислот[10], соединенных пептидной связью.
Образование пептидов происходит в результате реакции конденсации аминокислот (рис. 256). При взаимодействии аминогруппы одной аминокислоты с карбоксильной группой другой, между ними возникает ковалентная азот-углеродная связь, которую называют пептидной. В зависимости от количества аминокислотных остатков, входящих в состав пепти-
|
 Рис. 256. Образование дипептида.
Рис. 256. Образование дипептида.
На одном конце молекулы находится свободная аминогруппа (его называют N-концом), а на другом — свободная карбоксильная группа (его называют С-концом).
В широком смысле пептиды и белки — одно и то же. В более узком
— пептидами называют олигопептиды, содержащие до 10 остатков аминокислот, белками же называют полипептиды, состоящие из более чем 10 остатков.
| Структура белковой молекулы |
Выполнение белками определенных специфических функций зависит от пространственной конфигурации их молекул. Кроме того, клетке энергетически невыгодно держать белки в развернутой форме — в виде цепочки. Поэтому полипептидные цепи подвергаются укладке, приобретая определенную трехмерную структуру, или конформацию. Образование компактных конформаций возможно благодаря возникновению внутримолекулярных и межмолекулярных связей (прежде всего водородных), возникающих между различными группировками аминокислотных остатков полипептидных цепей, а также в результате гидрофобных взаимодействий между неполярными радикалами. Выделяют 4 уровня пространственной организации белков (рис. 257).
| Первичная структура |
Под первичной структурой белка понимают последовательность расположения аминокислотных остатков в одной или нескольких полипептидных цепях, составляющих молекулу белка.
Первым белком, у которого была выявлена аминокислотная последовательность, стал гормон инсулин. Исследования проводились в Кембриджском университете Ф.Сэнгером с 1944 по 1954 год. Было выявлено, что молекула инсулина состоит из двух полипептидных цепей (21 и 30 аминокислотных остатков), удерживаемых около друг друга дисульфидными мостиками. За свой кропотливый труд Ф.Сэнгер был удостоен Нобелевской премии.
|
 Рис. 257. Структура белковой молекулы:
1 — первичная; 2 — вторичная; 3 — третичная; 4 — четвертичная структуры.
Рис. 257. Структура белковой молекулы:
1 — первичная; 2 — вторичная; 3 — третичная; 4 — четвертичная структуры.
|
 Рис. 258. Водородные связи.
Рис. 258. Водородные связи.
| Вторичная структура |
Лишь незначительное количество белков имеет строго линейную структуру. Основная масса белков подвергается дальнейшей укладке, что приводит к образованию вторичной структуры белковой молекулы.
Вторичной структурой называют упорядоченное свертывание полипептидной цепи. Основным вариантом вторичной структуры является a-спираль, имеющая вид растянутой пружины. Она образована одной полипептидной цепью в результате возникновения внутримолекулярных водородных связей между
карбоксильными группами и аминогруппами, расположенными на соседних витках спирали (рис. 258). Практически все СО- и NН-группы принимают участие в образовании водородных связей. Они слабее пептидных, но, повторяясь многократно, придают данной конфигурации устойчивость и жесткость.
| Третичная структура |
Большинство полипептидных цепей приобретает вид компактной глобулы. Третичная структура — это способ укладки полипептидных цепей глобулярных белков, возникающий в результате образования изгибов, приводящих к наложению одних участков спирали на другие, и определенных взаимодействий между этими участками. При образовании третичной структуры происходит сшивание участков в результате возникновения химических связей (водородных, ионных, дисульфидных) и установления гидрофобных взаимодействий между боковыми цепями аминокислотных остатков.
|
 Рис. 259. Связи, стабилизирующие третичную структуру:
А — водородные, Б — ионные, В — дисульфидные.
Рис. 259. Связи, стабилизирующие третичную структуру:
А — водородные, Б — ионные, В — дисульфидные.
У некоторых белков третичная структура стабилизируется ди-
сульфидными ковалентными связями, возникающими между атомами серы двух остатков цистеина. Третичная структура специфична для каждого белка.
| Четвертичная структура |
Четвертичная структура характерна для сложных белков, молекулы которых образованы двумя и более глобулами. В одних белках субъединицы одинаковы или имеют сходное строение, в других различны. Однако они всегда образуют единое целое и располагаются в молекуле симметрично.
Субъединицы удерживаются в молекуле благодаря ионным, гидрофобным и электростатическим взаимодействиям. Иногда при образовании четвертичной структуры между субъединицами возникают дисульфидные связи.
Наиболее изученным белком, имеющим четвертичную структуру, является гемоглобин. Он образован двумя a-субъединицами (141 аминокислотный остаток) и двумя b-субъединицами (146 аминокислотных остатков). С каждой субъединицей связана молекула гема, содержащая железо.
| Классификация белков |
Обычно белки классифицируют по отдельно взятым признакам.
По химическому составу различают:
© простые — белки, состоящие только из аминокислот (фибрин, трипсин);
© сложные — белки, содержащие помимо аминокислот еще и небелковую — простетическую группу, которая может быть представлена ионами металлов (металлопротеины — гемоглобин), углеводами (гликопротеины), липидами (липопротеины), нуклеиновыми кислотами (нуклеопротеины).
По форме молекулы различают:
© Глобулярные — белки, имеющие сферическую форму — форму компактной глобулы (инсулин, белки крови, ферменты). Для них наиболее важна третичная структура. Хорошо растворимы в воде, в разбавленных водных растворах кислот, оснований и солей.
© Фибриллярные — белки, имеющие вытянутую форму молекул, обычно собранных в пучки, образующие волокна (кератин ногтей, волос, перьев, паутины, шелка, коллаген сухожилий). Для них наиболее важна вторичная структура. Нерастворимы в воде. Отличаются большой механической прочностью.
| Денатурация и ренатурация белков |
Внешние факторы (изменение температуры, солевого состава среды, рН, радиация) могут вызывать нарушение структурной организации молекулы белка. Процесс утраты трехмерной конформации, присущей данной молекуле белка, называют денатурацией (рис. 260). Причиной денатурации является разрыв связей, стабилизирующих определенную структуру белка. Причем первоначально рвутся наиболее слабые связи, а при ужесточении условий и более сильные. Поэтому сначала утрачивается четвертичная, затем третичная и вторичная структуры. Вместе с тем, денатурация не сопровождается разрушением полипептидной цепи. Изменение пространственной конфигурации приводит к изменению свойств белка и, как следствие, делает невозможным выполнение белком свойственных ему биологических функций.
|
 Рис. 260. Денатурация и ренатурация белка:
1 — молекула белка третичной структуры; 2 — денатурированный белок; 3 — восстановление третичной структуры в процессе ренатурации.
Рис. 260. Денатурация и ренатурация белка:
1 — молекула белка третичной структуры; 2 — денатурированный белок; 3 — восстановление третичной структуры в процессе ренатурации.
Денатурация может быть:
© Обратимой, если возможно восстановление свойственной белку структуры. Такой денатурации подвергаются, например, рецепторные белки мембраны.
© Необратимой, если восстановление пространственной конфигурации белка невозможно. Обычно это происходит при разрыве большого количества связей, например, при варке яиц.
Если белок подвергся обратимой денатурации, то при восстановлении нормальных условий среды он способен полностью восстановить свою структуру и, соответственно, свои свойства и функции. Процесс восстановления структуры белка после денатурации называется ренатурацией.
| Функции белков |
Благодаря сложности, разнообразию форм и состава, белки играют важную роль в жизнедеятельности клетки и организма в целом. Функции их разнообразны.
| Строительная (структурная) функция |
Одна из важнейших — строительная. Белки участвуют в образовании клеточных и внеклеточных структур: входят в состав клеточных мембран, шерсти, волос, сухожилий, стенок сосудов и т.д.
| Транспортная функция |
Некоторые белки способны присоединять различные вещества и переносить их к различным тканям и органам тела, из одного места клетки в другое. Например, белок крови гемоглобин присоединяет кислород и транспортирует его от легких ко всем тканям и органам, а от них в легкие переносит углекислый газ; в состав клеточных мембран входят особые белки, обеспечивают активный и строго избирательный перенос некоторых веществ и ионов из клетки во внешнюю среду и обратно.
| Регуляторная функция |
Большая группа белков организма принимает участие в регуляции процессов обмена веществ. Такими белками являются гормоны — биологически активные вещества, выделяющиеся в кровь железами внутренней секреции. Они влияют на активность ферментов, тем самым, замедляя или ускоряя обменные процессы, изменяют проницаемость клеточных мембран, поддерживают постоянство концентрации веществ в крови и клетках, участвуют в процессах роста, размножения и т.д. Например, гормон инсулин регулирует уровень сахара в крови путем повышения проницаемости клеточных мембран для глюкозы, способствует синтезу гликогена, увеличивает образование жиров из углеводов.
| Защитная функция |
В ответ на проникновение в организм чужеродных белков или микроорганизмов (антигенов) образуются особые белки — антитела, способные связывать и обезвреживать их. Синтез этих белков, называемых иммуноглобулинами, происходит в лимфоцитах. Причем, практически на любой антиген, с которым клетка и организм никогда не встречались, лимфоциты способны синтезировать антитела. Фибрин, образующийся из фибриногена, способствует остановке кровотечений.
| Двигательная функция |
Особые сократительные белки участвуют во всех видах движения клетки и организма: образовании псевдоподий, мерцании ресничек и биении жгутиков у простейших, сокращении мышц у многоклеточных животных, движении листьев у растений и др.
| Сигнальная функция |
Весьма важна для жизни клетки сигнальная функция белков. В поверхностную мембрану клетки встроены молекулы белков, способных изменять свою третичную структуру в ответ на действие факторов внешней среды. Так происходит прием сигналов из внешней среды и передача команд в клетку.
| Запасающая функция |
Благодаря белкам в организме могут откладываться про запас некоторые вещества. Например, при распаде гемоглобина железо не выводится из организма, а сохраняется в селезенке, образуя комплекс с белком ферритином. К запасным белкам относятся белки яйца, белки молока.
| Энергетическая функция |
Белки являются одним из источников энергии в клетке. При распаде 1 г белка до конечных продуктов выделяется 17,6 кДж. Сначала белки распадаются до аминокислот, а затем до конечных продуктов — воды, углекислого газа и аммиака. Однако в качестве источника энергии белки используются тогда, когда другие (углеводы и жиры) израсходованы.
| Каталитическая функция |
Одна из важнейших функций белков. В состав клеток входит большое количество веществ, химически мало активных. Тем не менее, все биохимические реакции протекают с огромной скоростью, благодаря участию в них биокатализаторов — ферментов — веществ белковой природы.
| Общая характеристика ферментов |
Как отмечалось ранее, большинство химических реакций в организме протекает с участием катализаторов — ферментов. Ферменты[11] — специфические белки, присутствующие во всех живых клетках и играющие роль биологических катализаторов.
Сходство ферментов и неорганических катализаторов заключается в том, что они:
© снижают энергию активации[12];
© не изменяют направления реакции, а лишь изменяют скорость ее протекания;
© в катализируемой реакции всегда затрачивается меньше энергии, чем в некатализируемой.
Но, поскольку ферменты являются белками, это придает им особые свойства:
© если неорганический катализатор может использоваться в разных типах реакций, то ферменты катализируют только одну реакцию или один вид реакции;
© большинство неорганических катализаторов ускоряют химические реакции при очень высоких температурах, имеют максимальную эффективность в сильнокислой или сильнощелочной среде, при высоких давлениях, а большинство ферментов активны при температурах 35-45˚С, физиологических значениях кислотности раствора и при нормальном атмосферном давлении;
© скорость ферментативных реакций в десятки тысяч (а иногда и в миллионы раз) выше скорости реакций, идущих с участием неорганических катализаторов. Например, пероксид водорода без катализаторов разлагается медленно: 2Н202 → 2Н20 + 02. В присутствии солей железа (катализатора) эта реакция идет несколько быстрее. Фермент каталаза за 1 сек. расщепляет 100 тыс. молекул Н202.
Известно более 2000 различных ферментов, представленных белками с высокой молекулярной массой, например каталаза (М=252000).
| Строение ферментов |
Несмотря на большое количество и разнообразие ферментов, все их по особенностям строения молекул можно разделить на две группы:
©
|
 Рис. 261. Строение фермента:
1 — первичная структура молекулы ферментного белка (черным цветом обозначены аминокислоты, входящие в состав активного центра фермента); 2 — схематическое изображение фермент-субстратного комплекса.
Рис. 261. Строение фермента:
1 — первичная структура молекулы ферментного белка (черным цветом обозначены аминокислоты, входящие в состав активного центра фермента); 2 — схематическое изображение фермент-субстратного комплекса.
© двухкомпонентные — сложные белки.
У двухкомпонентных ферментов, помимо белковой части, имеется добавочная группа небелковой природы — кофактор, например многие витамины.
В молекуле ферментов выделяют особую часть, представляющую собой уникальное сочетание нескольких аминокислотных остатков, располагающихся в определенной части белковой молекулы. Ее называют активным центром фермента. Именно эта часть фермента вступает в контакт с субстратом. Поскольку аминокислотные остатки, образующие каталитический центр, расположены в различных участках полипептидной цепи, он возникает только тогда, когда белковая молекула приобретает характерную для нее третич-
ную структуру (рис. 261). Если под влиянием каких-либо факторов происходит изменение третичной структуры фермента, то, как правило, это приводит к деформации каталитического центра и изменению ферментативной активности.
| Свойства ферментов |
Ферменты как биологические катализаторы белковой природы обладают рядом важных свойств:
© все ферменты — глобулярные белки;
© ферменты обладают строгой специфичностью, каждый фермент катализирует только одну реакцию (или тип реакции);
© высокая активность, обеспечивающая протекание ферментативных реакций с большой скоростью;
© активность ферментов зависит от условий, в которых протекает реакция.
| Механизм действия ферментов |
Ферментативные реакции протекают в несколько этапов:
© На первом этапе происходит образование фермент-субстратного комплекса за счет возникновения связей субстратного центра фермента с субстратом (или субстратами).
|
 Рис. 262. Соответствие фермента и субстрата:
1 — гипотеза "ключа и замка"; 2 — гипотеза "руки и перчатки".
Рис. 262. Соответствие фермента и субстрата:
1 — гипотеза "ключа и замка"; 2 — гипотеза "руки и перчатки".
В 1959 году Д. Кошланд выдвинул гипотезу,
по которой пространственное соответствие структуры субстрата и активного центра фермента создается лишь в момент их взаимодействия друг с другом. Эту гипотезу называют гипотезой "руки и перчатки" (гипотезой индуцированного соответствия) (рис. 262).
© На следующем этапе происходит сама химическая реакция и образуется продукт (продукты) этой реакции.
© На конечном этапе фермент-субстратный комплекс распадается на фермент и продукт (продукты) реакции.
Поскольку все ферменты являются белками, их активность наиболее высока при физиологически нормальных условиях:
© Большинство ферментов наиболее активно работает только при определенной температуре. При повышении температуры до некоторого значения (в среднем до 5О°С) каталитическая активность растет (на каждые 10°С скорость реакции повышается примерно в 2 раза). При температуре выше 50°С белок подвергается денатурации и активность фермента падает.
© Для каждого фермента существует оптимальное значение рН, при котором он проявляет максимальную активность.
| Фермент | Характер катализируемой реакции | рН |
| Пепсин | Гидролиз белка | 1,5-2,5 |
| Липаза | Гидролиз жиров | 4,7-5,0 |
Большинство ферментов имеет максимальную активность в зоне рН поблизости от нейтральной точки. В резко кислой или резко щелочной среде хорошо работают лишь некоторые ферменты.
© При увеличении количества субстрата скорость ферментативной реакции растет до тех пор, пока количество молекул субстрата не станет равным количеству молекул фермента. При дальнейшем увеличении количества субстрата скорость увеличиваться не будет, так как происходит насыщение активных центров фермента.
© Так же на скорость реакции влияет увеличение концентрации фермента, так как в единицу времени преобразованиям подвергается большее количество молекул субстрата.
Углеводы
Углеводы, или сахариды, — органические вещества, в состав которых входит углерод, кислород, водород. Углеводы составляют около 1% массы сухого вещества в животных клетках, а в клетках печени и мышц — до 5%. Наиболее богаты углеводами растительные клетки (до 90% сухой массы). Химический состав углеводов характеризуется их общей формулой Сm(Н2О)n, где m≥n. Количество атомов водорода в молекулах углеводов, как правило, в два раза больше атомов кислорода (то есть как в молекуле воды). Отсюда и название — углеводы.
Различают две группы углеводов:
© простые сахара;
© сложные сахара, образованные остатками простых сахаров.
| Простые углеводы |
Простые углеводы называют моносахаридами, так как они не гидролизуются. Общая формула простых сахаров — (СН2О)n, где n ≥ 3.В зависимости от числа атомов углерода в молекуле моносахаридов различают: триозы (3С), тетрозы (4С), пентозы (5С), гексозы (6С), гептозы (7С). В природе наиболее широко распространены пентозы и гексозы.
| Свойства моносахаридов |
© Низкая молекулярная масса;
© сладкий вкус;
© легко растворяются в воде;
© кристаллизуются;
© относятся к редуцирующим (восстанавливающим) сахарам.
Важнейшие моносахариды: из пентоз — дезоксирибоза и рибоза, входящие в состав ДНК, РНК и АТФ; из гексоз наиболее распространены глюкоза, фруктоза и галактоза (общая формула С6Н12О6).
Молекулы моносахаридов могут иметь вид прямолинейных цепочек или циклических структур (рис. 263). Для пентоз и гексоз — наиболее характерна именно циклическая структура, линейные молекулы встречаются очень редко. Молекулы дисахаридов и полисахаридов также образованы
|
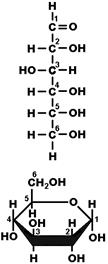 Рис. 263. Линейная и циклическая структура молекулы глюкозы.
Рис. 263. Линейная и циклическая структура молекулы глюкозы.
Моносахариды могут быть представлены в форме a- и b-изомеров (рис. 264). Гидроксильная группа при первом атоме углерода может располагаться как под плоскостью цикла (a-изомер), так и над ней (b-изомер). a- и b-изомеры играют важную роль, например, в образовании крахмала и целлюлозы.
|
 Рис. 264. Изомеры глюкозы:
1 — a-изомер; 2 — b-изомер.
Рис. 264. Изомеры глюкозы:
1 — a-изомер; 2 — b-изомер.
Наиболее важные моносахариды |
| Рибоза и дезоксирибоза |
|
 Рис. 265. Пентозы:
1 — рибоза; 2 — дезоксирибоза.
Рис. 265. Пентозы:
1 — рибоза; 2 — дезоксирибоза.
| Глюкоза (виноградный сахар) |
Одна из наиболее распространенных гексоз. В свободном виде встречается и у растений, и у животных. Глюкоза — это первичный источник энергии для клеток. Входит в состав важнейших ди- и полисахаридов. Обязательный компонент крови. Снижение ее количества приводит к немедленному нарушению жизнедеятельности нервных и мышечных клеток. Находясь в клетках, регулирует осмотическое давление.
| Фруктоза |
Широко распространена в природе. В свободном виде встречается в плодах. Особенно много ее в меде, фруктах. Значительно слаще глюкозы и других сахаров. Входит в состав олиго- и полисахаридов, участвует в поддержании тургора растительных клеток. Поскольку метаболизм фруктозы не регулируется инсулином, имеет важное значение при питании больных сахарным диабетом.
| Галактоза |
Пространственный изомер глюкозы. Входит в состав олигосахаридов, растительных и бактериальных полисахаридов. Вместе с глюкозой образуют важнейший дисахарид молока — лактозу, называемую молочным сахаром. Легко превращается в глюкозу.
| Сложные углеводы |
Сложными называют углеводы, молекулы которых при гидролизе распадаются с образованием простых углеводов. Их состав выражается общей формулой Сm(H2O)n, где m>n.
Среди сложных углеводов различают олигосахариды и полисахариды.
| Олигосахариды |
Олигосахаридами называют сахароподобные сложные углеводы, содержащие от 2 до 10 моносахаридных остатков.
В зависимости от количества остатков моносахаридов, входящих в молекулы олигосахаридов, различают дисахариды, трисахариды, тетрасахариды и т.д. Наиболее широко распространены в природе дисахариды.
| Свойства олигосахаридов |
© Сравнительно невысокая (несколько сотен) молекулярная масса;
© хорошая растворимость в воде;
© легко кристаллизуются;
© обладают, как правило, сладким вкусом;
© могут быть как редуцирующими, так и нередуцирующими.
| Дисахариды |
Дисахариды — олигосахариды, молекулы которых образованы двумя остатками моносахаридов. Встречаются в природе в свободном виде или в составе полисахаридов.
| Образование дисахаридов |
Дисахариды образуются в результате конденсации двух моносахаридов (чаще всего гексоз) (рис. 266). Связь, возникающую между двумя моносахаридами, называют гликозидной. Обычно она образуется между 1-м и 4-м
|
 Рис. 266. Образование дисахарида.
Рис. 266. Образование дисахарида.
| Сахароза (тростниковый сахар) |
Состоит из остатков глюкозы и фруктозы. Легко растворима в воде. Широко распространена в растениях. Углеводы, образовавшиеся в процессе фотосинтеза, в виде сахарозы оттекают из листьев. Сахароза легко превращается в крахмал и гликоген. Играет огромную роль в питании животных и человека. В основном сахарозу получают из сахарной свеклы и сахарного тростника.
| Лактоза (молочный сахар) |
Образована остатками глюкозы и галактозы. Плохо растворима в воде. Входит в состав молока. Является источником энергии для детенышей млекопитающих. В свободном виде обнаружена у некоторых растений. Используется в микробиологической промышленности для приготовления питательных сред.
| Мальтоза (солодовый сахар) |
Состоит из двух остатков глюкозы. Хорошо растворима в воде. Легко гидролизуется ферментом мальтаза с образованием двух молекул глюкозы.
| Полисахариды |
Высокомолекулярные органические вещества, биополимеры, мономерами которых являются простые углеводы. Чаще всего мономером полисахаридов является глюкоза, иногда манноза, галактоза и другие сахара. Как правило, в состав полисахаридов входит несколько сотен мономерных единиц.
| Свойства полисахаридов |
© Большая молекулярная масса (обычно сотни тысяч);
© не дают ясно оформленных кристаллов;
© либо нерастворимы в воде, либо образуют растворы, напоминающие по свойствам коллоидные;
© сладкий вкус не характерен;
© нередуцирующие углеводы.
| Образование полисахаридов |
Полисахариды образуются в результате реакции поликонденсации (рис. 267). Если в молекуле полисахарида присутствуют только 1,4-гликозидные связи, то образуется линейный, неразветвленный полимер (целлюлоза). Если присутствуют как 1,4, так и 1,6-гликозидные связи, полимер будет разветвленным (гликоген). 1,6-гликозидная связь образуется между остатками моносахаридов, входящих в состав разных линейных цепей.
|
 Рис.267. Образование разветвленного полисахарида.
Рис.267. Образование разветвленного полисахарида.
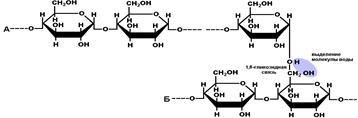
Наиболее важные полисахариды |
| Крахмал |
Основной резервный углевод растений. Общая формула (С6Н10О5)n, где n- количество остатков a-глюкозы. Нерастворим в холодной воде. В горячей воде образует раствор, по свойствам напоминающий коллоидный (крахмальный клейстер). Молекула крахмала примерно на 20% состоит из амилозы и на 80% из амилопектина. Линейные цепи амилозы состоят из нескольких тысяч остатков глюкозы и способны спирально свертываться, принимая более компактную форму. Амилопектин интенсивно ветвится, и за счет этого обеспечивается его компактность.
| Гликоген |
Основной резервный углевод животных и человека. Обнаружен также в грибах, дрожжах и зернах кукурузы. Содержится главным образом в печени (20%) и мышцах (4%). Служит источником глюкозы.
Молекула сходна с молекулой амилопектина, но сильнее ветвится. Гликоген сравнительно хорошо растворим в горячей воде.
| Целлюлоза (клетчатка) |
Основной структурный углевод клеточных стенок растений. Один из самых распространенных природных полимеров: в ней аккумулировано около 50% всего углерода биосферы. Целлюлоза нерастворима в воде, лишь набухает в ней. Является линейным полимером b-глюкозы. В отличие от крахмала, остатки глюкозы соединены в молекуле целлюлозы b-гликозидными связями, что исключает ее расщепление пищеварительными соками человека, так как у человека отсутствуют ферменты, способные разрывать b-гликозидные связи целлюлозы.
| Функции углеводов |
| Энергетическая |
Одна из основных функций углеводов. Углеводы — основные источники энергии в животном организме. Обеспечивают до 67% суточного энергопотребления (не менее 50%). При расщеплении 1 г углевода выделяется 17,6 кДж.
| Запасающая |
Запасающая функция выражается в накоплении крахмала клетками растений и гликогена клетками животных, которые играют роль источников глюкозы, легко высвобождая ее по мере необходимости.
| Опорно-строительная |
Углеводы входят в состав клеточных мембран и клеточных стенок (целлюлоза входит в состав клеточной стенки растений, из хитина образован панцирь членистоногих, различные олиго- и полисахариды образуют клеточную стенку бактерий). Продукты промежуточного обмена углеводов используются для синтеза липидов, аминокислот. Соединяясь с липидами и белками, образуют гликолипиды и гликопротеины. Рибоза и дезоксирибоза входят в состав мономеров нуклеотидов.
| Рецепторная |
Олигосахаридные фрагменты гликопротеинов и гликолипидов клеточных стенок выполняют рецепторную функцию, воспринимая сигналы, поступающие из внешней среды.
| Защитная |
Слизи, выделяемые различными железами, богаты углеводами и их производными (например, гликопротеинами). Они предохраняют пищевод, кишечник, желудок, бронхи от механических повреждений, препятствуют проникновению в организм бактерий и вирусов. Гепарин предотвращает свертывание крови в организме животных и человека.
Липиды
Липиды — сборная группа органических соединений, не имеющих единой химической характеристики. Их объединяет то, что все они являются производными высших жирных кислот, нерастворимы в воде, но хорошо растворимы в органических растворителях (эфире, хлороформе, бензине).
Липиды содержатся во всех клетках животных и растений. Содержание липидов в клетках составляет 5-15% сухой массы, но в жировой ткани может иногда достигать 90%.
В зависимости от особенности строения молекул различают:
© Простые липиды, представляющие собой двухкомпонентные вещества, являющиеся сложными эфирами высших жирных кислот и какого-либо спирта.
© Сложные липиды, имеющие многокомпонентные молекулы.
| Простые липиды Жиры |
Жиры широко распространены в природе. Они входят в состав организма человека, животных, растений, микробов, некоторых вирусов. Содержание жиров в биологических объектах, тканях и органах может достигать 90%.
Жиры — это сложные эфиры высших жирных кислот и трехатомного спирта — глицерина. В химии эту группу органических соединений принято называть триглицеридами. Триглицериды — самые распространенные в природе липиды.
| Жирные кислоты |
В составе триглицеридов обнаружено более 500 жирных кислот, молекулы которых имеют сходное строение. Как и аминокислоты, жирные кислоты имеют одинаковую для всех кислот группировку — карбоксильную группу (–СООН) и радикал, которым они отличаются друг от друга. Поэтому общая формула жирных кислот имеет вид R-CООН. Карбоксильная группа образует головку жирной кислоты. Она полярна, поэтому гидрофильна. Радикал представляет собой углеводородный хвост, отличающийся у разных жирных кислот количеством группировок –СН2. Он неполярен, поэтому гидрофобен. Большая часть жирных кислот содержит в "хвосте" четное число атомов углерода, от 14 до 22 (чаще всего 16 или 18). Кроме того, углеводородный хвост может содержать различное количество двойных связей. По наличию или отсутствию двойных связей в углеводородном хвосте различают:
© насыщенные жирные кислоты, не содержащие в углеводородном хвосте двойных связей;
© ненасыщенные жирные кислоты, имеющие двойные связи между атомами углерода (-СН=СН-).
| Образование молекулы триглицерида |
При образовании молекулы триглицерида каждая из трех гидроксильных (-ОН) групп глицерина вступает в реакцию
конденсации с жирной кислотой (рис. 268). В ходе реакции возникают три сложноэфирные связи, поэтому образовавшееся соединение называют сложным эфиром. Обычно в реакцию вступают все три гидроксильные группы глицерина, поэтому продукт реакции называется триглицеридом.
|
 Рис. 268. Образование молекулы триглицерида.
Рис. 268. Образование молекулы триглицерида.
Свойства триглицеридов |
Физические свойства зависят от состава их молекул. Если в триглицеридах преобладают насыщенные жирные кислоты, то они твердые (жиры), если ненасыщенные — жидкие (масла).
Плотность жиров ниже, чем у воды, поэтому в воде они всплывают и находятся на поверхности.
| Воски |
Воски — группа простых липидов, представляющих собой сложные эфиры высших жирных кислот и высших высокомолекулярных спиртов.
Воски встречаются как в животном, так и в растительном царстве, где выполняют главным образом защитные функции. У растений они, например, покрывают тонким слоем листья, стебли и плоды, предохраняя их от смачивания водой и проникновения микроорганизмов. От качества воскового покрытия зависят сроки хранения фруктов. Под покровом пчелиного воска хранится мед и развиваются личинки. Другие виды животного воска (ланолин) предохраняют волосы и кожу от действия воды.
| Сложные липиды Фосфолипиды |
Фосфолипиды — сложные эфиры многоатомных спиртов с высшими жирными кислотами, содержа-
|
 Рис. 269. Фосфолипид.
Рис. 269. Фосфолипид.
Как правило, в молекуле фосфолипидов имеется два остатка высших жирных и
один остаток фосфорной кислоты.
Фосфолипиды найдены и в животных, и в растительных организмах. Особенно много их в нервной ткани человека и позвоночных животных, много фосфолипидов в семенах растений, сердце и печени животных, яйцах птиц.
Фосфолипиды присутствуют во всех клетках живых существ, участвуя главным образом в формировании клеточных мембран.
| Гликолипиды |
Гликолипиды — это углеводные производные липидов. В состав их молекул наряду с многоатомным спиртом и высшими жирными кислотами входят также углеводы (обычно глюкоза или галактоза). Они локализованы преимущественно на наружной поверхности плазматической мембраны, где их углеводные компоненты входят в число других углеводов клеточной поверхности.
| Липоиды |
Липоиды — жироподобные вещества. К ним относятся стероиды (широко распространенный в животных тканях холестерин, эстрадиол и тестостерон — соответственно женский и мужской половые гормоны), терпены (эфирные масла, от которых зависит запах растений), гиббереллины (ростовые вещества растений), некоторые пигменты (хлорофилл, билирубин), часть витаминов (А, D, E, K ) и др.
| Функции липидов Энергетическая |
Основная функция липидов — энергетическая. Калорийность липидов выше, чем у углеводов. В ходе расщепления 1 г жиров до СО2 и Н2О освобождается 38,9 кДж. Единственной пищей новорожденных млекопитающих является молоко, энергоемкость которого определяется главным образом содержанием в нем жира.
| Структурная |
Липиды принимают участие в образовании клеточных мембран. В составе мембран находятся фосфолипиды, гликолипиды, липопротеины.
| Запасающая |
Жиры являются запасным веществом животных и растений. Это особенно важно для животных, впадающих в холодное время года в спячку или совершающих длительные переходы через местность, где нет источников питания (верблюды в пустыне). Семена многих растений содержат жир, необходимый для обеспечения энергией развивающееся растение.
| Терморегуляторная |
Жиры являются хорошими термоизоляторами вследствие плохой теплопроводимости. Они откладываются под кожей, образуя у некоторых животных толстые прослойки. Например, у китов слой подкожного жира достигает толщины 1 м. Это позволяет теплокровному животному обитать в холодной воде. Жировая ткань многих млекопитающих играет роль терморегулятора.
| Защитно-механическая |
Скапливаясь в подкожном слое, жиры не только предотвращают потери тепла, но и защищают организм от механических воздействий. Жировые капсулы внутренних органов, жировая прослойка брюшной полости обеспечивают фиксацию анатомического положения внутренних органов и защищают их от сотрясения, травмирования при внешних воздействиях.
| Каталитическая |
Эта функция связана с жирорастворимыми витаминами (А, D, E, K). Сами по себе витамины не обладают каталитической активностью. Но они являются кофакторами ферментов, без них ферменты не могут выполнять свои функции.
| Источник метаболический воды |
Одним из продуктов окисления жиров является вода. Эта метаболическая вода очень важна для обитателей пустынь. Так, жир, которым заполнен горб верблюда, служит в первую очередь не источником энергии, а источником воды (при окислении 1 кг жира выделяется 1,1 кг воды).
| Повышение плавучести |
Запасы жира повышают плавучесть водных животных.
Нуклеиновые кислоты
Нуклеиновые кислоты впервые были выделены Ф.Мишером в 1869 г. из ядер клеток гноя, а сам термин предложен А.Косселем в 1889 г.
К нуклеиновым кислотам относят высокополимерные соединения, распадающиеся при гидролизе на пуриновые и пиримидиновые основания, пентозу и фосфорную кислоту. Нуклеиновые кислоты содержат С, Н, О большое количество Р (8-10%) и N (15-16%).
Значение нуклеиновых кислот для живых организмов заключается в обеспечении хранения, реализации и передачи наследственной информации.
| Нуклеотиды |
Нуклеотид — мономер нуклеиновых кислот. Молекула нуклеотида состоит из трех частей: азотистого основания, пятиуглеродного сахара (пентозы) и фосфорной кислоты (рис. 270).
| Состав нуклеотидов Азотистые основания |
Азотистые основания являются главной частью нуклеотида. Они имеют циклическую структуру, в состав которой наряду с атомами углерода входят атомы других элементов, в частности азота. За присутствие в этих соединениях атомов азота они и получили название азотистых, а поскольку они обладают щелочными свойствами — оснований.
Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов.
Пиримидиновые основания являются производными гетероциклического соединения — пиримидина, имеющего в составе своей молекулы одно кольцо. К наиболее распространенным пиримидиновым основаниям относятся урацил, тимин, цитозин.
Пуриновые основания являются производными бициклического гетероцикла — пурина, имеющего два кольца: шестичленное и пятичленное.
К пуриновым основаниям относятся аденин и гуанин.
Во всех клетках — прокариотических и эукариотических — в состав нуклеиновых кислот входят эти пять основных азотистых оснований.
| Пятиуглеродный сахар |
Помимо азотистых оснований в образовании нуклеотидов принимают участие углеводный компонент, который представлен двумя сходными моносахаридами: рибозой или дезоксирибозой, относящихся к пентозам.
| Фосфорная кислота |
Третьим компонентом нуклеотидов является остаток фосфорной кислоты — фосфат. Именно наличие фосфата придает нуклеиновым кислотам свойства кислот.
|
 Рис. 270. Компоненты нуклеотидов:
1 — пятиуглеродный сахар; 2 — азотистые основания; 3 — фосфорная кислота.
Рис. 270. Компоненты нуклеотидов:
1 — пятиуглеродный сахар; 2 — азотистые основания; 3 — фосфорная кислота.
Образование нуклеотидов |
Как отмечалось ранее, нуклеотиды являются мономерами нуклеиновых кислот. Биосинтез нуклеотидов является первым этапом биосинтеза нуклеиновых кислот. Они их непосредственные предшественники.
Образование нуклеотида происходит в два этапа. На первом этапе в результате реакции конденсации образуется нуклеозид — комплекс азотистого основания с сахаром. На втором этапе нуклеозид подвергается фосфорилированию. При этом между остатком сахара и фосфорной кислотой возникает фосфоэфирная связь. Таким образом, нуклеотид представляет собой нуклеозид, соединенный с остатком фосфорной кислоты (рис. 271).
|
 Рис. 271. Образование нуклеотида.
Рис. 271. Образование нуклеотида.
Названия нуклеотидов отличаются от названий соответствующих оснований. И те, и другие принято обозначать заглавными буквами:
Таблица 6.
Виды нуклеотидов:
| Азотистое основание | Нуклеотид | Обозначение |
| Аденин | Адениловый | А |
| Гуанин | Гуаниловый | Г или G |
| Цитозин | Цитидиловый | Ц или С |
| Тимин | Тимидиловый | Т |
| Урацил | Уридиловый | У или U |
| Функции нуклеотидов |
Нуклеотиды являются мономерами, из которых построены полимерные цепи нуклеиновых кислот, они входят в состав важных коферментов (НАД, НАДФ, ФАД, КоА).
| Образование ди- и полинуклеотидов |
Динуклеотид представляет собой соединение, состоящее из остатков двух нуклеотидов. При конденсации двух нуклеотидов между 3'-углеродом остатка сахара одного нуклеотида и остатком фосфорной кислоты другого возникает сложноэфирная связь. Таким образом, остатки сахаров двух
|
 Рис.272. Образование динуклеотида
Рис.272. Образование динуклеотида
Возникновение фосфодиэфирных мостиков между 3' и 5'-углеродами остатков сахаров может происходить многократно. В результате образуются неразветвленные полинуклеотидные цепи. Один конец полинуклеотидной цепи заканчивается 5'-углеродом (его называют 5'-концом), другой –3'-углеродом (3'-концом).
| Полинуклеотиды |
В зависимости от углеводного компонента нуклеотидов, различают два класса нуклеиновых кислот:
© рибонуклеиновые кислоты (РНК), содержащие рибозу;
© дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу.
Нуклеотиды ДНК называют дезоксирибонуклеотидами, РНК — рибонуклеотидами.
| Дезоксирибонуклеиновая кислота |
Молекулы ДНК являются полимерами, мономерами которых являются дезоксирибонуклеотиды, образованные:
© остатком пятиуглеродного сахара — дезоксирибозы;
© остатком одного из азотистых оснований:
¨ пуриновых — аденина, гуанина;
¨ пиримидиновых — тимина, цитозина;
© остатком фосфорной кислоты.
| Структура молекулы ДНК |
ДНК представляет собой двойную спираль. Ее молекула образована двумя полинуклеотидными цепями, спирально закрученными друг около друга, и вместе вокруг воображаемой оси (рис. 273). Цепи ДНК антипараллельны (разнонаправлены), то есть против 3'-конца одной цепи находится 5'-конец другой. На периферию молекулы обращен сахаро-фосфатный остов, образованный чередующимися остатками дезоксирибозы и фосфатными группами. Внутрь молекулы обращены азотистые основания.
Диаметр двойной спирали ДНК — 2 нм, шаг общей спирали, на который приходится 10 пар нуклеотидов — 3,4 нм. Длина молекулы — до нескольких десятков и даже сотен микрометров. Молекулярный вес составляет десятки и сотни миллионов (для двойной спирали). В ядре клетки человека общая длина ДНК около 2м.

Трехмерная модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. американским биологом Дж.Уотсоном и английским физиком Ф.Криком (рис. 274). За свои исследования они были удостоены Нобелевской премии.
Полинуклеотидные цепи в молекуле ДНК удерживаются друг около друга благодаря возникновению водородных связей между азотистыми основаниями. Спаривание нуклеотидов не случайно, в его основе лежит принцип комплементарного взаимодействия пар оснований: против аденина одной цепи всегда располагается тимин на другой цепи, а против гуанина одной цепи — всегда цитозин другой, то есть аденин комплементарен тимину, а гуанин — цитозину (рис. 275). Комплементарностью называют способность нуклеотидов к избирательному соединению друг с другом.
|
 Рис. 275. Спаривание азотистых оснований.
Рис. 275. Спаривание азотистых оснований.
Комплементарность обеспечивается:
© взаимодополнением пространственных конфигураций молекул азотистых оснований;
© количеством водородных связей, возникающих между азотистыми основаниями (три водородные связи между гуанином и цитозином и две водородные связи между аденином и тимином).
Комплементарность пуриновых и пиримидиновых азотистых оснований обеспечивает одинаковое по всей длине двойной спирали расстояние между цепями.
Э.Чаргафф, обследовав огромное количество образцов тканей и органов различных организмов, выявил следующую закономерность: в любом фрагменте ДНК содержание остатков гуанина всегда точно соответствует содержанию цитозина, а аденина — тимину. Это положение получило название "правила Чаргаффа":
А + Г
А = Т; Г = Ц или ——— = 1
Ц + Т
Дж.Уотсон и Ф.Крик воспользовались этим правилом при построении модели молекулы ДНК.
Последовательность нуклеотидов одной цепи определяет последовательность нуклеотидов другой, поэтому две цепи молекулы ДНК комплементарны друг другу.
| Самоудвоение молекулы ДНК |
Одним из уникальных свойств молекулы ДНК является ее способность к самоудвоению — воспроизведению точных копий исходной молекулы. Благодаря этой способности молекулы ДНК, осуществляется передача наследственной информации от материнской клетки дочерним во время деления. Процесс самоудвоения молекулы ДНК называют репликацией.
Репликация — сложный процесс, идущий с участием ферментов (ДНК-полимераз) (рис. 276). Репликация осуществляется полуконсервативным способом, то есть под действием ферментов молекула ДНК раскручивается и около каждой цепи, выступающей в роли матрицы, по принципу комплементарности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Раскручивание молекулы происходит на небольшом отрезке (несколько десятков нуклеотидов), называемом репликативной вилкой. После окончания синтеза дочерних цепей ДНК на данном участке и соединения их с материнскими раскручивается новый отрезок, и цикл репликации повторяется. Таким образом, репликативная вилка перемещается вдоль молекулы, пока не дойдет до точки окончания синтеза.
|
 Рис. 276. Репликация ДНК.
Рис. 276. Репликация ДНК.
Во время репликации энергия молекул АТФ не расходуется, так как для синтеза дочерних цепей при репликации используются не дезоксирибонуклеотиды (содержат один остаток фосфорной кислоты), а дезоксирибонуклеозидтрифосфаты (содержат три остатка фосфорной кислоты). При включении дезоксирибонуклеозидтрифосфатов в полинуклеотидную цепь два концевых остатка отщепляются, и освободившаяся энергия используется на образование сложноэфирной связи между нуклеотидами.
Рибонуклеиновые кислоты
Молекулы РНК являются полимерами, мономерами которых являются рибонуклеотиды, образованные:
¨ остатком пятиуглеродного сахара — рибозы;
¨ остатком одного из азотистых оснований:
¨ пуриновых — аденина, гуанина;
¨ пиримидиновых — урацил, цитозина;
¨ остатком фосфорной кислоты.
| Структурная организация РНК |
Молекула РНК представляет собой неразветвленный полинуклеотид, имеющий третичную структуру. В отличие от ДНК, она образована не двумя, а одной полинуклеотидной цепочкой. Однако ее нуклеотиды также способны образовывать водородные связи между собой, но это внутри–, а не межцепочечные соединения комплементарных нуклеотидов. Цепи РНК значительно короче цепей ДНК.
Информация о структуре молекулы РНК заложена в молекулах ДНК. Синтез молекул РНК происходит на матрице ДНК с участием ферментов РНК-полимераз и называется транскрипцией. Последовательность нуклеотидов в РНК комплементарна кодирующей цепи ДНК и идентична, за исключением замены тимина на урацил, некодирующей цепи.
Если содержание ДНК в клетке относительно постоянно, то содержание РНК сильно колеблется. Наибольшее количество РНК в клетках наблюдается во время синтеза белка.
Существует три основных класса рибонуклеиновых кислот:
© информационная (матричная) РНК — иРНК;
© транспортная РНК — тРНК;
© рибосомальная РНК — рРНК.
| Информационная РНК |
Наиболее разнообразный по размерам и стабильности класс. Все они являются переносчиками генетической информации из ядра в цитоплазму. Они служат матрицей для синтеза молекулы белка, т.к. определяют аминокислотную последовательность первичной структуры белковой молекулы.
На долю иРНК приходится до 5% от общего содержания РНК в клетке.
| Транспортная РНК |
Молекулы транспортных РНК содержат обычно 75-86 нуклеотидов. Молекулярная масса молекул тРНК » 25000. Молекулы тРНК играют роль посредников в биосинтезе белка — они доставляют аминокислоты к месту синтеза белка, в рибосомы. В
|
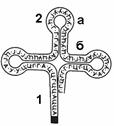 Рис. 277. Строение тРНК:
1 — акцепторное плечо; 2 — антикодоновое плечо (а — петля; б — "стебель").
Рис. 277. Строение тРНК:
1 — акцепторное плечо; 2 — антикодоновое плечо (а — петля; б — "стебель").
Каждый вид тРНК имеет характерную только для него последовательность нуклеотидов. Однако у всех молекул имеется несколько внутримолекулярных комплементарных участков, благодаря наличию которых все тРНК имеют третичную структуру, напоминающую по форме лист клевера (рис. 277).
Молекулы всех тРНК имеют четыре основных плеча:
© акцепторное;
© антикодоновое;
© два боковых.
Каждое плечо состоит из "стебля", образованного комплементарными парами оснований, и петель из неспаренных оснований.
Акцепторное плечо через 3¢-гидроксильную группу аденозильного остатка связывает тРНК с аминокислотой. Антикодоновое плечо содержит триплет нуклеотидов (антикодон), комплементарный кодону иРНК.
| Рибосомная РНК |
На долю рибосомальной РНК (рРНК) приходится 80-85% от общего содержания РНК в клетке. Рибосомная РНК состоит из 3-5 тыс. нуклеотидов. В комплексе с рибосомными белками рРНК образует рибосомы — органеллы, на которых происходит синтез белка.
Основное значение рРНК состоит в том, что она обеспечивает первоначальное связывание иРНК и рибосомы и формирует активный центр рибосомы, в котором происходит образование пептидных связей между аминокислотами в процессе синтеза полипептидной цепи.
Дата добавления: 2018-09-20; просмотров: 462; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!