Плауновидные - Lycopod di yta.
Лекция №5-2
Систематика растений. Высшие споровые растения (Моховидные, Плауновидные, Хвощевидные, Папоротниковидные).
Высшие споровые растения.

Высшие растения.
У большинства высших растений тело дифференцировано на органы - корень, стебель и листья, состоящие из хорошо обособленных тканей. В жизненном цикле высших растений четко выражено чередование спорофита (2n) и гаметофита (n). Органы полового размножения многоклеточные. Женский - архегоний - состоит из расширенной нижней части - брюшка, где формируется яйцеклетка, и верхней суженной - шейки, раскрывающейся при созревании яйцеклетки. Мужской орган полового размножения - антеридий - имеет вид мешка, внутри которого образуется множество сперматозоидов. У голосеменных антеридии подверглись редукции, а у покрытосеменных редуцированы и антеридии, и архегоний. Из зиготы у высших растений образуется зародыш - зачаток спорофита.
Царство объединяет не менее 300000 ныне существующих и вымерших видов, дошедших до нас лишь в ископаемом состоянии. Известные науке растения обычно делят на 9 отделов:
- Риниевые — Rhyniophyta.-вымерли
- Зостерофилловые — Zostero- phyllophyta.-вымерли
- Моховидные — Bryophyta.
- Плауновидные — Lycopodiophyta.
- Псилотовидные — Psilotophyta.
- Хвощевидные — Equiseto phyta,
- Папоротниковидные — Polypodiophyta.
- Голосемянные — Gymno- spermae, или Pinophyta.
- Покрытосемянные, или цветковые — Angiospermae, или Magnolio phyta.
|
|
|
Два первых отдела растений — риниевые и зостерофилловые давно и полностью вымерли. Остальные отделы наряду с вымершими включают и ныне существующие виды. Представители всех перечисленных отделов, исключая моховидных, характеризуются преобладанием спорофита, в органах которого имеются сосуды и (или) трахеиды, поэтому их нередко называют сосудистыми растениями (Tracheo phyta).
Растения делят на две, очень неравные по числу современных видов группы — споровые и семенные растения. У споровых процессы споро- и гаметогенеза разобщены во времени и пространстве: спорофиты, образующие споры, и гаметофиты, продуцирующие гаметы, являются самостоятельными организмами, ведущими раздельное существование. К споровым относятся все отделы растений, кроме голосемянных и покрытосемянных, которые называют семенными растениями. У семенных растений споро- и гаметогенез тесно сопряжены: редуцированный женский гаметофит развивается на спорофите, не покидая его, а мужской гаметофит (функционально выполняющий роль гаметы) переносится к женским половым органам в виде пыльцевого зерна. Зигота у семенных растений развивается в зародыш, окруженный специальными покровами, — так называемое семя. Размножаются эти растения семенами. По времени происхождения споровые значительно древнее семенных растений. Они были первыми растениями, завоевавшими сушу. Семенные растения произошли от споровых в процессе эволюции.
|
|
|
Споровые растения.
Моховидные - Bryophyta.
Общее число видов около 35 тыс.
В подавляющем большинстве моховидные — низкорослые многолетние растения размером от 1 мм до нескольких сантиметров, реже до 60 см и более. Тело у части моховидных представляет собой слоевище, а у других расчленено на стебель и листья. Характерный признак всех моховидных — отсутствие корней. Всасывание воды и прикрепление к субстрату у них осуществляют ризоиды, представляющие собой выросты эпидермы. Моховидные могут быть однодомными или двудомными. Внутреннее строение их относительно простое. У листостебельных форм ассимиляционная, механическая и проводящие ткани более или менее обособлены. Элементы проводящих тканей сходны с трахеидами и ситовидными трубками. Особенно своеобразен цикл развития моховидных. Как и для всех растений, для них характерно правильное чередование полового и бесполого поколений. Однако доминирует в цикле развития гаплоидный гаметофит, что резко отличает мхи от всех остальных растений. Другая особенность этой группы в отличие от других споровых состоит в том, что гаметофит и спорофит представляют собой как бы одно растение. Бесполое поколение (спорофит) у моховидных нередко называется спорогоном и представлено небольшой коробочкой со спорами и ножкой, нижняя часть которой превращена в гаусторий (присоску), внедряющийся в тело гаметофита. Спорофит, таким образом, лишен самостоятельности, по существу, паразитирует на нем.
|
|
|
Развитие полового поколения моховидных начинается с момента прорастания споры. Прежде всего, развивается ветвистое нитчатое (у большинства мхов) или пластинчатое (у сфагнума) многоклеточное образование — протонема (предросток), на которой закладываются почки. У одних мхов из почек вырастают пластинчатые слоевища, у других — облиственные стебли — гаметофоры, на которых развиваются половые органы моховидных.
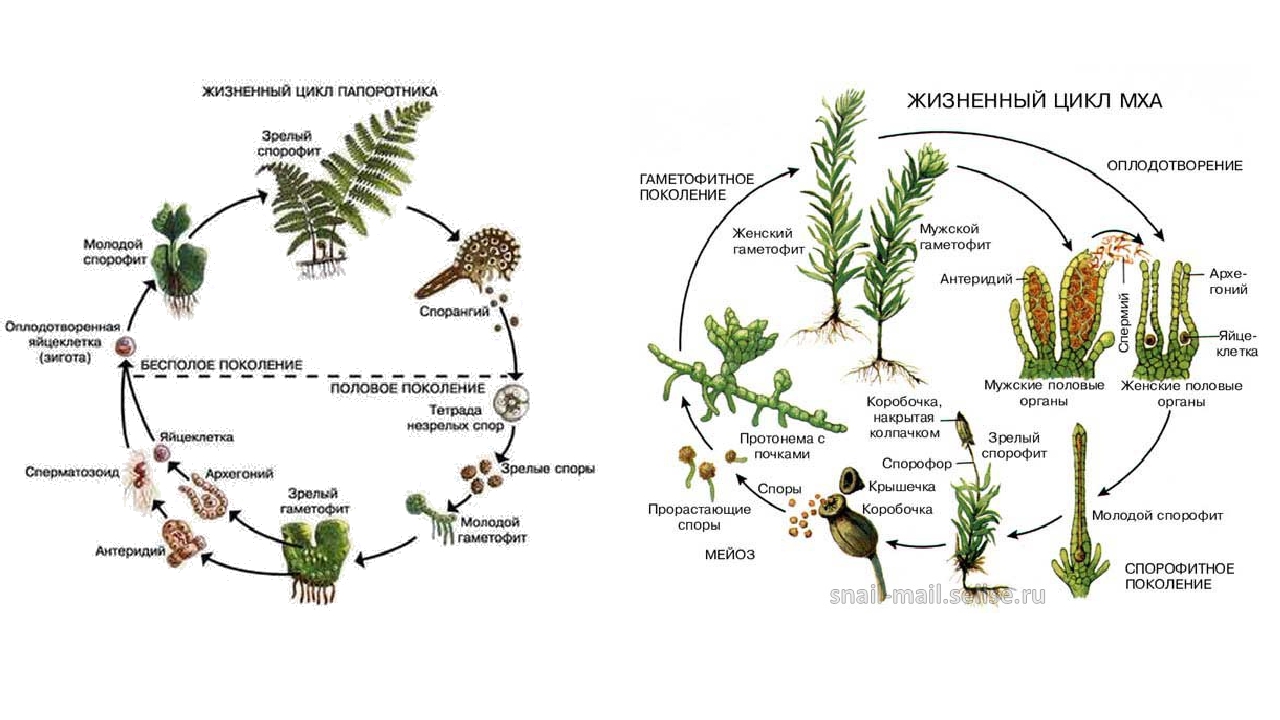
Строение. В жизненном цикле моховидных, как и других высших растений, имеется чередование двух фаз: спорофита и гаметофита. Однако доминирует (преобладает) гаметофит, в то время как у всех других высших растений доминирует спорофит. Именно поэтому моховидные рассматривают как самостоятельную боковую ветвь в эволюции высших растений.
|
|
|
Гаметофит обычно многолетний. Он представляет собой листовидный таллом или растение в виде побега, расчлененного на стебель и листья. Корней нет, их функцию выполняют ризоиды - выросты поверхностных клеток тела. Органы полового размножения многоклеточные. Спорофит, называемый у моховидных спорогоном, играет подчиненную роль. Он представляет собой цилиндрическую ножку, заканчивающуюся шаровидной, эллиптической или цилиндрической коробочкой, внутри которой образуется спорангий со спорами. Спорогон паразитирует на гаметофите, так как получает от него воду и необходимую пищу. Максимальная длина тела (гаметофита и спорогона) 60 см.
Моховидные по своей организации и экологии еще близки к водорослям. Как и у водорослей, у них нет сосудов и корней. Некоторые примитивные представители имеют вегетативное тело в виде стелющегося таллома с верхушечным (дихотомическим) ветвлением, похожего на таллом водорослей. Оплодотворение связано с водой. Среди моховидных, как и среди водорослей, нет одревесневающих форм.
Распространение. Моховидные распространены на всех континентах мира, но неравномерно. В тропических странах - преимущественно в горах. Незначительное число видов растет в засушливых условиях, например в степях. Некоторые виды ведут эпифитный образ жизни на корке деревьев или водный. Основное же разнообразие видов сосредоточено во влажных местах Северного полушария, в областях с умеренным и холодным климатом. В сложении растительного покрова, особенно тундр, болот и лесов, им принадлежит важная роль.
Классификация. Моховидные подразделяют на три класса: Антоцеротовые, Печеночники, Листостебельные мхи. Наибольшее значение имеют два последних класса.

Класс Печеночники - Hepaticopsida.
Общее число видов около 10 тыс. Распространелы повсеместно. Примитивность строения тела печеночников свидетельствует об их древности.
Маршанция обыкновенная (Marchantia polymorpha) - типичный представитель класса. Гаметофит в виде пластинчатого таллома, длиной 10 - 12 см, ветвление верхушечное. С обеих сторон он покрыт эпидермой. Верхняя эпидерма имеет вентиляционные отверстия - устьица. Они окружены специальными клетками, расположенными в четыре ряда. Под устьицами имеются воздушные камеры. Нижняя эпидерма дает выросты - одноклеточные ризоиды и красноватые или зеленоватые чешуйки, которые иногда принимают за редуцированные листья. Под верхней эпидермой расположена ассимиляционная ткань, состоящая из вертикальных столбиков паренхимных клеток с хлоропластами. Ниже находится слой тонкостенных бесхлорофилльных паренхимных клеток. Следовательно, таллом маршанции имеет дорсивентральное строение.
На верхней стороне таллома образуются особые веточки - подставки, а на них - органы полового размножения. Маршанция - двудомное растение. На одних экземплярах подставки имеют форму сидящей на ножке девятилучевои звездочки, между лучами которой на нижней стороне расположены архегонии. На других - подставки имеют форму сидящего на ножке восьмиугольного щитка, на верхней стороне которого расположены антеридии, погруженные в антеридиальные полости. В брюшке архегония образуется яйцеклетка. После ее слияния со сперматозоидом из зиготы образуется спорогон. Он представляет собой коробочку на короткой ножке, которая прикреплена к гаметофиту гаусторией. Внутри коробочки из спорогенных клеток в результате мейоза образуются гаплоидные споры, а также элатеры - мертвые удлиненные клетки со спирально утолщенной стенкой, служащие для разрыхления массы спор, а также для выбрасывания их из коробочки. В благоприятных условиях из споры развивается предросток, или протонема. Это небольшая нить. Из ее верхушечной клетки вырастает таллом маршанции.
Вегетативное размножение осуществляется выводковыми тельцами линзовидной формы, имеющими зеленый цвет. Они образуются на верхней стороне таллома в особых корзиночках в результате деления клеток, выстилающих их дно.
Виды маршанции имеют широкое распространение. Наиболее часто их можно встретить во влажных местах: на берегах озер и рек, по оврагам и в травянистом покрове под пологом леса.
Класс Листостебельные мхи - Bryopsida.
Общее число видов около 25 тыс. Многие виды распространены в приполярных странах Северного полушария. На огромных территориях в тундре, на болотах, в лесах они доминируют в растительном покрове, значительно влияя на обеспеченность суши влагой.
Гаметофит представляет собой прямостоячую стеблевидную ось - каулидий, покрытую листовидными выростами - филлидиями. Условно их можно называть стебель и листья. На нижней части стебля образуются многоклеточные ризоиды (не у всех). Ветвление боковое. Нарастание осей происходит в результате деления пирамидальной верхушечной клетки. Оно может быть моноподиальным или симподиальным. В соответствии с этим органы полового размножения и спорогон размещаются на вершине гаметофита или на боковых разветвлениях.
Класс подразделяют на три подкласса: Андреевые мхи, Сфагновые мхи, Бриевые (Зеленые) мхи. Наибольшее значение имеют два последних подкласса.
Подкласс Сфагновые мхи - Sphaqnidae.
Сфагновые мхи имеют довольно однообразное строение и поэтому трудны для определения. Гаметофит их представляет собой сильноветвящееся, особенно в верхней части, растение. Разветвления густо покрыты листьями. Сфагновые мхи живут в очень влажной среде. В связи с этим у них нет ризоидов и влага поступает непосредственно в стебель, который при основании с течением времени отмирает. Строение стебля несложное. В центре его находится сердцевина из тонкостенных паренхимных клеток, выполняющих проводящую и запасающую функции. Она окружена корой, состоящей из двух слоев: склеродермы, выполняющей механическую функцию, и гиалодермы, выполняющей водозапасающую функцию. Клетки гиалодермы крупные, мертвые, стенки их имеют круглые отверстия, через которые полости смежных клеток сообщаются друг с другом, а также с внешней средой. Иногда эти клетки несут спиральные утолщения. Лист состоит из одного ряда клеток, резко различающихся как по структуре, так и по выполняемой функции. Одни из них живые, хлорофиллоносные, другие - мертвые, относительно более крупные, со спирально утолщенными стенками, пронизанные отверстиями, по строению сходные с водозапасающими клетками гиалодермы, их называют гиалиновыми. Гиалиновые клетки способны накапливать и долго удерживать огромное количество воды, в 30 - 40 раз превышающее массу самого растения.
Гаметофиты - однодомные и двудомные. Антеридии образуются в пазухах листьев на разветвлениях стебля. Около них листья окрашены в красноватый цвет. Архегонии на укороченных разветвлениях. В результате слияния сперматозоида с яйцеклеткой возникает зигота, которая представляет собой начало диплоидной фазы - спорогона. Спорогон состоит из ножки и коробочки. Ножка сильно укорочена, луковицеобразная, но ко времени созревания спор верхушка стебля гаметофита сильно вырастает и выносит коробочку кверху (ложная ножка). В центре коробочки помещается округлая колонка, над которой размещен в виде свода спорангий со спорогенной тканью. Стенка коробочки прочная, многослойная. Наружный хлорофиллоносный слой содержит большое число недоразвитых устьиц. Коробочка имеет крышечку, которая во время созревания спор отскакивает, и споры рассеиваются. Элатер нет. Из спор образуется сначала зеленая пластинчатая протонема, а затем из почек, расположенных на ней, - взрослый гаметофит, который и доминирует в жизненном цикле.
Структура сфагновых отличается примитивностью: пластинчатая протонема, отсутствие проводящего пучка и ризоидов, слабая дифференциация коробочки.
Значение сфагновых в природе очень велико. Накапливая огромное количество воды и разрастаясь плотными дернинами, они вызывают заболачивание огромных пространств, достигающих зоны тундры. Для осушения их осуществляют агромелиоративные работы. С другой стороны, старые болота имеют важное хозяйственное значение для разработки залежей торфа. Нарастание пласта торфа в наиболее благоприятных условиях происходит медленно - слой толщиной 1 см образуется примерно за 10 лет.
Подкласс Бриевые (зеленые) мхи - Bryidae.
Число видов 24,6 тыс. Распространены более широко, чем сфагновые мхи. Живут в разнообразных экологических условиях от тундры и лесотундры до степей и пустынь. Наиболее типичные местообитания бриевых мхов, где они доминируют или формируют сплошной покров, - тундра, болота и некоторые типы лесов. Каждому местообитанию свойственны свои виды. Бриевые мхи по сравнению со сфагновыми отличаются большим разнообразием строения. Органы полового размножения закладываются у одних видов на главной оси, у других - на боковых. У некоторых видов ветвление не выражено.
Политрих обыкновенный, кукушкин лен (Polytrichum commune) - это один из распространенных представителей бриевых мхов. Он растет в лесу, на полянах, окраинах болот.
Стебель гаметофита прямостоячий, неветвистый, высотой 15 см и более, густо покрыт листьями. Подземная часть его простирается в почве почти горизонтально, на ней образуются ризоиды. В центре стебля находится концентрический проводящий пучок, состоящий из вытянутых клеток, сходных с трахеидами и ситовидными трубками. Он окружен паренхимой, также выполняющей проводящую функцию. С наружной стороны паренхима граничит со склеродермой (корой). Внешний слой ее, состоящий из бесцветных клеток, называют гиалодермой.
Листья расположены по спирали. Они состоят из линейной пластинки с заостренной зубчатой верхушкой и пленчатого влагалища. На верхней стороне листа расположены ассимиляционные пластинки. Жилка с механическими и проводящими элементами расширена.
Гаметофит двудомный. Архегонии бутылковидной формы расположены на верхушке женского гаметофита, антеридии мешковидной формы - на верхушке мужского. Между архегониями и антеридиями имеются стерильные нити - парафизы. После оплодотворения из зиготы образуется спорогон, состоящий из длинной ножки и коробочки. Коробочка прямостоячая или более или менее косо расположенная, призматическая, четырех-пятиграниая, покрыта ржаво-войлочным колпачком, образующимся из стенок архегония. Коробочка состоит из урночки и крышечки. Нижняя часть урночки сужена в шейку. На границе урночки и шейки в эпидерме имеются устьица. В центре урночки расположена колонка, которая у крышечки расширяется и формирует эпифрагму - тонкостенную перегородку, закрывающую урночку. Вокруг колонки расположен спорангий в виде цилиндрического мешка, прикрепленного к стенке и колонке особыми нитевидными образованиями. Урночка имеет специальное приспособление для рассеивания спор - перистом, представляющий собой ряд зубчиков с тупыми верхушками, расположенных по краю урночки. Между зубцами, способными к гигроскопическим движениям, и эпифрагмой имеются отверстия, через которые в сухую погоду высыпаются споры. Из споры вырастает протонема в виде зеленой ветвящейся нити. На ней формируются почки, из которых со временем развиваются взрослые гаметофиты.
Плауновидные - Lycopod di yta.
Плауновидные - одни из наиболее древних высших растений. Они представляют мелколистную линию эволюции. Современные виды - многолетние вечнозеленые травянистые растения, среди вымерших были и древесные формы.
Современные плауновидные — это многолетние травянистые растения с простыми листьями и дихотомическим ветвлением Стебель хорошо развит и имеет спиральное, супротивное или мутовчатое листорасположение. На подземных корневищах обычно образуются придаточные корни. Верхушечная меристема со временем теряет свою активность, поэтому плауновидные ограничены в росте.
Спорофиллы по форме, размерам и цвету похожи на обычные вегетативные листья или отличаются от них. Чередуясь с ассимиляционными листьями, они образуют на стебле спороносные зоны или собраны в расположенные на верхушках ветвей стробилы, нередко называемые спороносными колосками. Среди плауновидных имеются равно- и разноспоровые представители. Листья современных плауновидных энационного типа.

Строение. Спорофит имеет надземный побег с мелкими, иногда чешуевидными листьями (микрофиллами). Они слабо дифференцированы, имеют 1 - 2 неветвящиеся жилки. Узлы и междоузлия выражены слабо. Подземная часть спорофита представлена корневищем с придаточными корнями. Ветвление надземных и подземных осей верхушечное. Спорангии располагаются на верхней стороне листьев (спорофиллов), собранных на концах осей в колоски, реже образующих на стебле спороносные зоны. Споры одинаковой или разной величины.

Гаметофиты подземные, длиной 2 - 20 мм, питаются сапрофитно. Оплодотворение связано с водой.
Классификация. Отдел делят на два класса: Плауновые и Полушниковые.
Класс Плауновые - Lycopodiopsida.
Спорофиты - травянистые многолетние растения. Стебель и корень не имеют камбия. Листья без язычков. Споры одинаковой величины. Гаметофиты обоеполые, созревают в течение 1 - 15 лет. Многие виды вымерли. В современной флоре класс представлен двумя родами. Наиболее многочисленный и широко распространенный из них - род плаун (Lycopodium). Хозяйственное значение плаунов невелико. Животные их не поедают. Плауны служат сырьем для производства лекарств. Издавна используют споры плаунов, содержащие невысыхающее масло. Их применяют в качестве детской присыпки, а иногда при фасонном литье для обсыпания стенок моделей, чтобы отливаемая деталь имела гладкие стенки и легко отделялась от формы.
Плаун булавовидный (Lycopodium clavatum). Растение, широко распространенное в хвойных лесах.
Спорофит представлен длинным ползучим побегом с вертикальными ответвлениями и придаточными корнями. Побег и корни имеют верхушечное ветвление. Проводящий пучок расположен в центре стебля. Центральный цилиндр занимает небольшую часть. Широкая зона коры пронизана листовыми следами. Камбия нет. На поверхности стебля и листьев имеется эпидерма с устьичными аппаратами. Листовая пластинка линейная, цельнокрайняя, заканчивается длинным тонким волоском.
Спороносные колоски, венчающие вертикальные побеги, расположены на довольно длинных ножках по два (реже по 3 - 5). Колосок цилиндрической формы, состоит из оси, на которой плотно сидят спорофиллы - чешуевидные треугольные листья с заостренными и загнутыми кверху верхушками. На верхней стороне спорофилла расположен на короткой ножке почковидный спорангий со спорами. Споры одинаковые, мелкие, тетраэдрической формы. Спородерма имеет два слоя: наружный - экзину, и внутренний - интину. Спорангий растрескивается поперечной щелью.
Споры падают на землю, и на глубине нескольких сантиметров из них медленно, в течение 12 - 15 лет, развивается гаметофит. По форме он напоминает луковичку, позднее разрастается и становится блюдцевидным диаметром до 2 см. Гаметофит бесцветный. Клетки, расположенные под эпидермой, находятся в симбиозе с мицелием гриба. У некоторых видов гаметофит образуется на поверхности почвы, и тогда в его клетках появляются хлоропласты. Антеридии и архегонии размещены на верхней стороне и погружены в паренхимную ткань. Сперматозоиды многочисленные, двухжгутиковые. Оплодотворение связано с водой. Зигота не имеет периода покоя, из нее сразу же образуется зародыш спорофита. Он вначале внедрен в ткань гаметофита и в какой-то мере питается за его счет, но вскоре корни его проникают в почву, и начинается долгая самостоятельная жизнь спорофита.
Класс Полушниковые - Isoetopsida.
Спорофиты представлены деревьями, имеющими вторичное утолщение стебля, и многолетними травами, отчасти сохранившими способность к вторичному утолщению. Древесные виды полностью вымерли. Листья на поверхности, обращенной к стеблю, имеют небольшой вырост - язычок. Споры разной величины. Гаметофиты раздельнополые, созревают в течение нескольких недель. Наиболее многочисленным и широко распространенным является род селагинелла (Selaginella). Используют лишь немногие ее виды как лекарственные и декоративные растения.
Селагинелла селаговидная (Selaginella selaginoides). В нашей стране изредка растет на влажных субальпийских лугах, расположенных в горах европейской части. Спорофит по внешнему виду похож на спорофит плауна. В центре стебля находится проводящий пучок, подвешенный на нитях из паренхимных клеток. Эпидерма стебля без устьичных аппаратов. Листья одинаковые по форме и величине, язычок со временем отваливается. В клетках мезофилла имеется по 1 -2 пластинчатых хлоропласта. Клетки эпидермы также содержат хлоропласты. Устьичные аппараты размещаются чаще на нижней стороне листа. Спороносные колоски расположены на верхушке побегов. Спорофиллы заметно отличаются от вегетативных листьев, они несут язычки, которые не отваливаются. В пазухах спорофиллов каждого колоска сидят на коротких ножках мега- и микроспорангии. В мегаспорангии образуется четыре мегаспоры, в микроспорангии - множество микроспор. Освободившись из спорангиев, мега- и микроспоры в благоприятных условиях прорастают. Внутри микроспоры образуется мужской гаметофит. Он состоит из одной вегетативной клетки и одного крайне редуцированного антеридия, внутри которого образуются двухжгутиковые сперматозоиды. Женский гаметофит также почти целиком формируется внутри мегаспоры. Разрастаясь, гаметофит разрывает покровы мегаспоры, Через трехлучевую щель выдвигается часть тела гаметофита, на которой образуются архегонии и ризоиды. Оплодотворение связано с водой. Из зиготы вырастает зародыш.
Таким образом, разноспоровость обусловила разнополость гаметофитов и их редукцию. Это сыграло важнейшую роль в эволюции растений.
Хвощевидные - Equisetophyta.
Древовидные представители полностью вымерли, в современной флоре сохранились только травянистые.
Все современные хвощи — многолетние корневищные травы с мутовками бурых редуцированных листьев, утративших хлорофилл. Проводящие элементы ксилемы представлены различного типа трахеидами, а иногда и сосудами. Флоэма состоит из ситовидных элементов и паренхимных клеток. Спорангии у хвощей располагаются на спорангиофорах, собранных на верхушках стеблей в стробилы, находится воротничок, представляющей собой редуцированное листовое влагалище. Спорангиофоры в виде шестиугольных щитков на ножках размещены на оси стробила мутовками. На внутренней стороне щитка располагаются 4-16 вытянутых вдоль ножки спорангиев. При созревании спор щитки подсыхают и раздвигаются, наружная стенка спорангия при этом легко разрушается, и споры рассеиваются ветром. Из внешнего слоя оболочки споры при созревании формируются спирально обернутые вокруг ее тела ленты - элатеры, способные отгибаться и совершать гигроскопические движения.
Все современные хвощевидные - равноспоровые растения. Их одно- или обоеполый гаметофит (заросток) представлен очень маленьким, величиной в несколько миллиметров, зеленым наземным растением. В антеридиях образуются сперматозоиды с большим числом ундулиподиев. Оплодотворение происходит в присутствии капельно-жидкой воды, после чего из зиготы без периода покоя начинает развиваться новый спорофит. Все хвощи обладают глубоко залегающими, сильно разветвленными корневищами, в узлах которых нередко развиваются клубеньки, богатые крахмалом. От узлов корневищ во множестве отходят придаточные корни.
Строение. Характерная особенность спорофита - боковое ветвление с мутовчатым расположением боковых побегов. Листья также собраны в мутовки. Узлы и междоузлия четко выражены. При основании междоузлий часто имеется интеркалярная меристема. Листья редуцированные, от средних до мелких, с одной средней жилкой. Спорангии расположены на спорангиофорах - гомологах спорофиллов, но отличающихся от них структурой. Выражен гетероталлизм (физиологическая разноспоровость) - из одинаковых по размеру спор образуются разнополые гаметофиты. Спородерма, кроме интины и экзины, имеет еще наружный слой - перину. Она состоит из двух спирально закрученных лент - элатер, прикрепленных к экзине. Элатеры выполняют двойную функцию: распространения спор и соединения спор в группы, что обеспечивает совместное произрастание мужского и женского гаметофитов.

Гаметофиты мелкие, величиной в несколько миллиметров, зеленые, одно- или (реже) обоеполые. Оплодотворение связано с водой. Зародыш не имеет периода покоя.
Классификация. Отдел подразделяют на четыре класса: Гиениевые, Клинолистные, Каламитовые, Хвощовые. Первые три класса вымерли. Хвощовые представлены в современной флоре обоих полушарий.
Класс Хвощовые- Equisetopsida
Класс включает один род - хвощ (Equisetum). Общее число видов 30 - 35, на территории бывшего СССР - 13. Распространены по всему земному шару, кроме Австралии. Растут преимущественно в условиях влажных местообитаний. По внешнему облику хвощи близки к своим древесным предкам - каламитовым, но значительно меньше их. Надземные побеги у хвощей, как правило, однолетние, лишь у немногих видов - многолетние, вечнозеленые. Отдельные виды имеют важное значение как кормовые растения (хвощ ветвистый - Е. ratnosissimuin, хвощ пятнистый - Е. variegatiun, хвощ зимующий - Е. hiemale). Однако у большинства видов стенки клеток эпидермы инкрустированы кремнеземом, что обесценивает их кормовое значение. Многие виды - трудно искореняемые сорняки пастбищ и полей, особенно с кислыми почвами (хвощ полевой- Е. arvense). Некоторые виды ядовиты для домашних животных (хвощ болотный - Е. palustre, хвощ дубравный - Е. nemorosuin). Хвощи используют как лекарственные растения. Стебли их употребляют вместо наждачной бумаги.
Хвощ полевой (Equisetum arvense) - один из наиболее широко распространенных видов. Это многолетнее травянистое растение. Растет на полях и залежах как сорняк. Его подземная часть представлена корневищем, проникающим в почву на глубину до 1 м. Некоторые укороченные боковые разветвления корневища превращаются в клубни, заполненные запасным крахмалом. В узлах мутовками расположены листовые влагалища и придаточные корни. Корень имеет первичное строение, в коре находятся крупные воздухоносные полости. Надземные побеги двух видов: спороносные, возникающие ранней весной, и стерильные, образующиеся позже и вегетирующие до поздней осени. У других видов хвоща надземные побеги одинаковы. Стерильный побег мутовчато-разветвленный, зеленый, ребристый, в узлах несет сросшиеся в трубку листовые влагалища, заканчивающиеся черными с белой каймой зубцами. Зубцы представляют собой редуцированные листовые пластинки. Органом фотосинтеза служит стебель. На поверхности он имеет однослойную эпидерму с устьичными аппаратами. Под эпидермой расположена кора, состоящая из участков механической и ассимиляционной ткани. Под ними лежит слой основной паренхимы, пронизанный крупными полостями. Заканчивается первичная кора эндодермой. Центральный цилиндр в центре имеет сердцевину из основной паренхимы, клетки которой со временем расходятся, образуя полости. Проводящие ткани собраны в пучки, расположенные по периферии центрального цилиндра. Пучки коллатеральные, закрытые, стебель, как и корень, не способен к вторичному утолщению.
Спороносные побеги более толстые, бурые, без хлорофилла, неветвистые, высотой 15 - 30 см. Они также охвачены в узлах трубчатыми влагалищами с 8 - 9 крупными зубцами. Спороносные колоски образуются на верхушке. После спороношения эти побеги отмирают. Спорангиофор состоит из шестигранного щитка, ножки, прикрепляющей щиток к оси колоска, мешковидных спорангиев, расположенных по нижнему краю щитка. Споры одинаковой величины. Элатеры имеют вид спирально закрученных лент с ложковидными расширениями на концах. На почве из спор вырастают хлорофиллоносные гаметофиты в виде лопастных пластинок, физиологически различные. Одни из них - мужские, с антеридиями, формирующими многожгутиковые сперматозоиды, другие - женские, с архегониями. Оплодотворение связано с водой. Зародыш спорофита не имеет периода покоя.
Таким образом, морфологическая равноспоровость у этого вида сочетается с физиологической разноспоровостыю.
Дата добавления: 2021-12-10; просмотров: 52; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!