ДЕМЭКОЛОГИЯ - ЭКОЛОГИЯ ПОПУЛЯЦИЙ
Понятие о популяции
Любой вид животного, растения, микроорганизма представляет собой сложную биологическую систему, важнейшими элементами которой являются внутривидовые группировки - популяции. Вид - это таксономическая и экологическая единица, а популяция рассматривается в качестве структурной единицы вида и единицы эволюции.
Каждый вид (совокупность всех особей этого вида) занимает определенную территорию - ареал, в различных частях которого наблюдаются разные условия. Часто группировки особей, располагающиеся в разных частях ареала вида, настолько изолированы друг от друга, что не могут контактировать, и не скрещиваются. Число этих группировок зависит от численности и исторического (филогенетического) возраста вида, от размеров ареала и от других причин. Группировки особей с общим генофондом, сходной морфологией, единым жизненным циклом и представляют собой популяции.
Термин «популяция» происходит от латинского слова populus (народ) и в дословном переводе означает «население». Понятие «популяция» является одним из центральных в биологии, а генетические, эволюционные и экологические подходы к изучению популяции объединяются в особое направление - популяционную биологию, разделом которой является популяционная экология, или демэкология.
Популяцией в экологии называют группу особей одного вида, находящихся во взаимодействии между собой и совместно населяющих общую территорию. Расширенное понятие о популяции дается в следующих определениях:
«Под популяцией понимается совокупность особей определенного вида, в течение достаточно длительного времени (большого числа поколений) населяющая определенное пространство, внутри которого практически осуществляется та или иная степень панмиксии и нет заметных изоляционных барьеров; которая отделена от соседних таких же совокупностей особей данного вида той или иной степенью давления тех или иных форм изоляции» (Н.В. Тимофеев-Ресовский и др., 1973).
«Популяция - это элементарная группировка организмов определенного вида, обладающая всеми необходимыми условиями для поддержания своей численности необозримо длительное время в постоянно изменяющихся условиях среды» (С.С. Шварц, 1967).
«Популяция - это любая, способная к самовоспроизведению совокупность особей одного вида, более или менее изолированная в пространстве и времени от других аналогичных совокупностей того же вида» (А.М. Гиляров, 1990).
«Популяция - это группировка особей одного вида, населяющих определенную территорию и характеризующихся общностью морфобиологического типа, специфичностью генофонда и системой устойчивых функциональных взаимосвязей» (И.А. Шилов, 1988).
Популяция как группа совместно обитающих особей одного вида является первой надорганизменной биологической макросистемой. С позиции системной иерархии к экологическим системам относятся надорганизменные - от видовых популяций до многовидовых сообществ и биосферы.
Основным свойством экологических систем, в том числе и популяций, является то, что они не статичны, а находятся в беспрерывном изменении, в движении, которое существенно отражается на структурно-функциональной организованности, продуктивности, биологическом разнообразии и устойчивости системы. Популяционный уровень занимает особое место в системе организации живой материи.
С одной стороны, популяция является элементарной единицей биоценотического взаимодействия, входя в функционально-экологический ряд различных уровней организации жизни: организм - популяция - биоценоз - биогеоценоз - биосфера.
С другой стороны, популяция является элементарной единицей эволюционного процесса, включаясь в генетико-эволюционный ряд, отражающий филогенетические связи таксонов разного уровня: организм - популяция - вид - род - семейство - отряд - класс - царство (рис. 5).
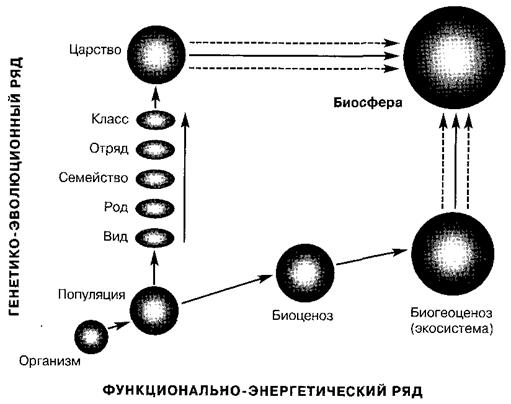
Рис. 5. Положение популяции в структуре биологических систем биосферы
(по И.А. Шилову, 1988)
Популяционная структура видов
Виду и популяциям свойственна структурированность. Вид, как правило, включает множество популяций. Изоляция между ними почти никогда не бывает абсолютной: между отдельными популяциями происходит обмен особями благодаря миграциям. Степень изолированности популяций зависит от способности к расселению, от наличия географических преград в пределах ареала вида (широкие реки, проливы, горные хребты и т.п.), а также от характера местообитаний.
Популяционная структура вида подробно рассмотрена в работах Н.П. Наумова, С.С. Шварца, К.М. Завадского и др. Широкое распространение в экологии получила концепция иерархии популяций в зависимости от размеров занимаемой ими территории. Профессор Московского университета Н.П. Наумов предложил классификацию популяций на ландшафтно-биотопической основе, выделяя элементарную, экологическую и географическую популяции (рис. 6).

Рис. 6. Пространственные подразделения популяций
Элементарная (локальная) популяция - это совокупность особей, занимающих какой-то небольшой участок однородной площади. Число элементарных популяций, на которые распадается вид, зависит от разнородности условий в биогеоценозе: чем они однообразнее, тем меньше элементарных популяций и наоборот. Нередко смешение особей элементарных популяций, происходящее в природе, стирает границы между ними.
Экологическая популяция - это население одного типа местообитания (биотопа), характеризующееся общим ритмом биологических циклов и характером образа жизни. Это наиболее мелкие территориальные группировки, которые формируются как совокупность элементарных популяций. Например, белка (Sciurus vulgaris) заселяет различные типы леса. Поэтому могут быть выделены «сосновые», «елово-пихтовые» и другие ее экологические популяции. Они слабо изолированы друг от друга, и обмен генетической информацией между ними происходит довольно часто, но реже, чем между элементарными популяциями.
Географическая популяция - совокупность особей одного вида, населяющих территорию с однородными условиями существования и обладающих общим морфологическим типом и единым ритмом жизненных явлений и динамики населения. Географические популяции относительно изолированы. Они различаются размерами особей, плодовитостью, рядом экологических, физиологических, поведенческих и других особенностей. Примером разных географических популяций могут служить популяции белки в заенисейской тайге и смешанных лесах Смоленщины, а также степная и тундровая популяции узкочерепной полевки.
Под влиянием ряда факторов географическая популяция может приобретать устойчивые особенности, отчетливо выделяющие ее из соседних, такую популяцию называют географической расой или подвидом (рис. 7). Вид белки обыкновенной, например, насчитывает более 20 подвидов.
В природе границы и размеры популяций определяются не только особенностями заселяемой территории, но и, главным образом, свойствами самой популяции. В основе всего лежит степень ее генетического и экологического единства. Раздробление вида на множество мелких территориальных группировок является процессом приспособления к величайшему разнообразию местных условий. Это увеличивает генетическое многообразие вида, обогащая его генофонд.
 |
Рис. 7. Популяционная структура вида
Лекция 7.
Структура популяции
Структура популяции - это определенная организация, формирующаяся, с одной стороны, на основе биологических свойств вида, а с другой - под влиянием абиотических факторов среды и популяций других видов. Структура популяции не стабильна.
Различают пространственную (территориальную), возрастную, половую, генетическую, поведенческую и другие виды структуры популяции.
Пространственная структура популяции - это особенность размещения особей популяции в пространстве. Она зависит как от свойств мест обитания, так и от биологических особенностей вида. Она может изменяться во времени, зависит от сезона года, от численности популяции и т.д. Пространственное размещение особей в популяциях может быть равномерным, случайным и групповым (рис. 8).
При равномерном (регулярном) распределении особи размещены через более или менее равные промежутки, например, деревья в зрелом сосновом лесу. В действительности же равномерное распределение особей встречается в природе редко.
При случайном (диффузионном) распределении особи размещены неравномерно и их встречи друг с другом носят случайный характер. Такой тип распределения широко представлен среди растений и многих видов животных. Случайное расселение имеет место в однородной среде.
При групповом (мозаичном) расселении особи встречаются группами, как, например, осоковые кочки на низинном болоте, стада млекопитающих или колонии птиц. Групповое размещение обеспечивает популяции более высокую устойчивость по отношению к неблагоприятным условиям.

Рис. 8. Пространственное размещение особей в популяциях
На больших географических пространствах со временем особи одной и той же популяции при распределении могут образовывать комбинации из этих трех типов размещения. Примером может служить расселение деревьев в лесу. Вначале происходит групповое расселение, а затем размещение может становиться равномерным (сосновые леса). Расселение тли и колорадского жука вначале может быть случайным, а по мере их размножения приобретает групповой или равномерный характер.
Границы популяции могут быть или хорошо выражены, когда территория разделена препятствием, таким, как горный хребет или река, или смазаны. Многие животные, ведущие подвижный образ жизни, расселяются активно. Это приводит к интенсивному перемешиванию популяций и стирает границы между ними. Например, очень подвижные и активно перемещающиеся песцы, некоторые птицы и другие животные имеют огромные ареалы без резких границ между популяциями. У малоподвижных и пассивно перемещающихся организмов (земноводных, наземных моллюсков), наоборот, популяции четко разграничены даже на сравнительно небольшой территории. Территория, занимаемая популяцией прыткой ящерицы (Lacerta agilis), может колебаться от 0,1 до нескольких гектаров. Популяция северного оленя занимает территорию более 100 кв. км.
Для животных как подвижных организмов ведущее значение в определении характера пространственной структуры популяции имеет степень привязанности к территории. По типу использования пространства все подвижные животные делятся на две группы: оседлые и кочевые (номадные), кроме этого существует ряд промежуточных групп.
Видам, для которых характерен оседлый образ жизни, как правило, свойственен интенсивный тип использования территории, при котором отдельные особи или их группировки в течение длительного времени эксплуатируют ресурсы на относительно ограниченном пространстве. Для видов, отличающихся кочевым образом жизни, характерен экстенсивный тип использования территории, при котором ресурсы потребляются обычно группами особей, постоянно перемещающимися в пределах обширной площади.
Распространение организмов за пределы популяции называется расселением. Способы расселения отражают, каким образом популяция занимает все большее пространство. Среди них выделяют следующие: анемохория (распространение ветром), гидрохория (водой), фитохория (растениями), зоохория (животными) и антропохория (человеком). Расселение может быть пассивным (аллохория) и активным (автохория).
Передвижения животных, вызванные изменением условий существования или связанные с циклами их развития, называются миграциями. Они могут быть регулярными (суточными и сезонными) и нерегулярными (при засухах, наводнениях, пожарах и т.д.). Классический пример сезонной миграции - перелеты птиц. Нерегулярные миграции, как правило, носят хаотический характер в отличие от большей части организованных, регулярных.
Миграции свойственны непаразитирующим животным. Распространение паразитирующих организмов связано с инвазиями. Инвазия (от лат. invasio - нападение, вторжение) - это заражение растений, животных и человека паразитами животного происхождения. Источниками инвазии являются организмы - носители паразитов, а также вода и корм. Инвазии приводят к вспышке численности паразитирующих организмов (простейших, червей, клещей и некоторых членистоногих) и поражению большого числа организмов - хозяев. Среди инвазионных заболеваний человека и животных наиболее известны гельминтозы (возбудители - черви), акарозы (возбудители - клещи), антомозы (возбудители - насекомые), а также малярия, лейшманиоз, амёбиоз, таксоплазмоз и другие (возбудители - простейшие).
Относительная территориальная изоляция популяции приводит к изоляции воспроизводства - ограничению свободы скрещивания. Популяция распадается на несколько панмиктических подразделений - близкородственных объединений особей по полу и возрасту, живущих в непосредственной близости друг от друга и потому часто контактирующих между собой. Такими микропопуляциями у растений являются парцеллы, а у животных - демы (стада, колонии и др.), которые существуют в течение жизни одного или нескольких поколений.
Половая структура популяции - это количественное соотношение особей по полу. Соотношение полов популяции устанавливается по генетическим законам, а затем на него влияет среда. У большинства видов пол будущей особи определяется в момент оплодотворения в результате перекомбинации половых хромосом. Сцепленные с полом признаки часто определяют различия в морфологии (размеры, окраска), физиологии (темпы роста, сроки полового созревания), экологии и поведении самцов и самок. Например, самцы комаров семейства Culicidae, в отличие от кровососущих самок, в имагинальный период либо не питаются совсем, либо ограничиваются слизыванием росы, потреблением нектара растений.
Соотношение особей по полу, особенно размножающихся самок в популяции, имеет большое значение для дальнейшего роста ее численности. В природе есть виды, отличающиеся высокой смертностью самок (например, ондатры, пингвины, летучие мыши), и наоборот, более высокой смертностью самцов (большие синицы, фазаны, многие грызуны). Так, у некоторых летучих мышей доля самок в популяции после зимней спячки снижается до 20% .
На соотношение полов в популяции влияют условия среды. У некоторых видов пол изначально определяется не генетическими, а экологическими факторами. Например, у растений Arisaema japonica пол зависит от накопления запасов питательных веществ в клубнях. Из крупных клубней вырастают экземпляры с женскими цветками, из мелких - с мужскими. У рыжих лесных муравьев (Formica rufa) из яиц, отложенных при температуре ниже +20°С развиваются самцы, а при более высокой температуре - самки. Механизм этого явления заключается в том, что мускулатура семяприемника, где хранится после копуляции сперма, активизируется лишь при высоких температурах, обеспечивая оплодотворение откладываемых яиц. Из неоплодотворенных яиц у перепончатокрылых развиваются лишь самцы.
В пределах ареала видов половая структура популяций растений более или менее постоянна, однако изменение внешних условий меняет соотношения полов. Например, в засушливый 1975 год в Зауралье резко уменьшилось число женских форм у шалфея степного (в 10 раз).
Генетическая структура популяции характеризуется разной степенью генетического разнообразия особей. Совокупность генов, которые имеются у особей конкретной популяции, называется генофондом. Совокупность всех генов, сосредоточенных в хромосомах одного организма, называется генотипом. С позиций генетики, популяция - совокупность генотипов. Если соотношение генотипов в популяции неизменно в поколениях, то популяция устойчива, в ней существует генотипическое равновесие. Несмотря на изменчивость в ее структурных частях, популяция как целостная система устойчиво сохраняет генофонд, наследованный от предковой популяции.
Генотип, взаимодействуя с условиями среды, формирует фенотип. Фенотипом называются элементарные признаки организма, определяющие индивидуальные особенности его строения и жизнедеятельности (морфологические, физиологические и поведенческие), которые зависят от взаимодействия генотипа с условиями среды.
Совокупность внешних признаков организма, отражающих его приспособленность к условиям среды, образует жизненную форму или биоморфу. Существование в течение многих поколений особей в популяции с двумя и более отчетливо выраженными формами, различающимися строением и выполняемыми функциями, называется полиморфизмом. Полиморфными видами, например, являются береза бородавчатая (Betula pendula), ель европейская (Picea abies), сосна обыкновенная (Pinus sylvestris).
У ели европейской три формы ветвления (гребенчатая, компактная и плоская лапчатая), шишки красного, зеленого и пурпурного цвета. В ельниках можно встретить деревья с гладкой, чешуйчатой, пластинчатой и ольховидной корой. Различают формы ели и по строению шишек (тупо-чешуйчатые и острочешуйчатые). Причем ель с острочешуйчатыми шишками растет быстрее и имеет древесину лучшего качества.
Универсальным свойством всего живого, от вирусов и микроорганизмов до высших растений и животных, является способность давать мутации - внезапные естественные или вызываемые искусственно, наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма. Мутации лежат в основе наследственной изменчивости и проявляются в фенотипе взрослого организма в результате того, что они изменяют процессы его онтогенеза. Мутационная изменчивость не отвечает условиям окружающей среды и, как правило, нарушает существующие адаптации.
Ведущим фактором видообразования является индивидуальный естественный отбор - выживание особей, наиболее приспособленных к среде обитания. В процессе борьбы за существование проявляется биологическая разнокачественность составляющих популяцию особей. Часть особей гибнет, другие выживают и размножаются. Вновь возникающие мутации комбинируются с генами, уже прошедшими отбор особей, их фенотипы меняются, и на этой основе возникают новые адаптации, вызывающие дальнейшее преобразование организмов.
Относительная пространственная обособленность популяции приводит к ее репродуктивной изоляции - ограничению свободы скрещивания особей разных популяций. Эта изоляция обеспечивает уникальность генофонда популяции и возможность ее самостоятельной эволюции. Однако в природе не существует полностью изолированных популяций и может происходить миграция (отток и приток) генов, приводящих к изменениям их генетической структуры.
Возрастная структура популяции определяет все возрастные группы особей, в том числе всех стадий и фаз развития организма, например, личинки и куколки насекомых, проростки растений. В популяции животных выделяют три экологических возраста: предрепродуктивный (довоспроизводящий), репродуктивный (воспроизводящий) и пострепродуктивный (послевоспроизводящий). Длительность каждого возраста по отношению к общей продолжительности жизни не одинакова у разных видов. У современного человека на каждую возрастную группу приходится примерно треть жизни. У многих животных и растений наиболее длительным бывает предрепродуктивный период. У поденок (Ephemeroptera), например, он продолжается несколько лет, у разных видов цикад (Cicadidea) - 13 и 17 лет (длительное развитие личинок). Репродуктивный же возраст их составляет всего несколько дней (период размножения взрослых особей), пострепродуктивный - практически отсутствует. Популяции, у особей которых короткий предрепродуктивный период, быстро восстанавливают свою численность, например полевки (Clethrionomys).
В жизненном цикле растений выделяют около десяти возрастных состояний, объединенных в четыре периода (таблица 4).
Возрастное состояние особи - это этап ее онтогенеза, на котором она характеризуется определенными отношениями со средой. Для наглядности приведем несколько примеров:
- проростки имеют смешанное питание за счет запасных веществ семени и собственной ассимиляции;
- ювенильные растения переходят к самостоятельному питанию, у них уже отсутствуют семядоли;
- у имматурных растений начинается ветвление побега;
- переход растений в генеративный период определяется не только появлением цветков и плодов, но и внутренней биохимической и физиологической перестройкой организма.
Таблица 4 - Возрастные периоды и состояния у семенных растений
| Период | Возрастное состояние | Индекс |
| I. Латентный (период покоя) | 1. Семена | sm |
| II. Прегенеративный (вегетативный) | 2. Проросток (всход) 3. Ювенильное 4. Имматурное 5. Виргинильное | Pi j im v |
| III. Генеративный (плодоносящий) | 6. Молодое 7. Зрелое 8. Старое | g1 g2 gз |
| IV. Постгенеративный (старческий) | 9. Субсенильное 10.Сенильне 11.Отмирающее | ss s sc |
Соотношение особей популяции по этим состояниям называют возрастным спектром популяции. Он отражает количественные соотношения разных возрастных групп. Возрастной спектр связан с интенсивностью смертности и рождаемости организмов. Возрастная структура популяции может изменяться под действием внешних факторов, так как они контролируют процессы и рождаемости, и смертности.

Рис. 9. Возрастная структура популяций: 1 - инвазионная (растущая); 2 - нормальная (стабильная); 3 - регрессивная (сокращающаяся)
По возрастному спектру оценивается способность популяции к самоподдержанию и ее устойчивость к внешним воздействиям: чем сложнее возрастной спектр, тем устойчивее воспроизводство популяции. Наибольшей жизнеспособностью отличаются популяции, в которых все возрасты представлены относительно равномерно. Такие популяции называются нормальными. Если в популяции преобладают старческие особи, это свидетельствует о наличии отрицательных факторов, нарушающих репродуктивные функции. Такие популяции называются регрессивными, или вымирающими. Популяции, представленные в основном молодыми особями, рассматриваются как внедряющиеся, или инвазионные. Жизненность их не вызывает опасений, но велика вероятность вспышек чрезмерно высокой численности особей. Особенно опасно, если такие популяции представлены видами, которые ранее здесь отсутствовали.
Для сопоставления численности особей разных возрастов в популяциях строятся гистограммы возрастных структур (рис. 9).
Исследования возрастной структуры популяций растений и животных редки из-за чрезмерной сложности и трудоемкости количественного учета особей разных возрастных групп. Исключение составляют популяции человека. Периодически проводимые переписи, позволяют достаточно полно охарактеризовать возрастной состав населения, который отражается в специальных диаграммах (рис. 10).

Рис. 10. Возрастная пирамида населения России
Изучение распределения организмов по возрастам имеет большое значение в охотничьем и рыбном промыслах, в рубке леса и т.д. Если взять, например, охотничий промысел, то преобладание молодых особей свидетельствует о благополучном размножении вида и позволяет ожидать в следующем сезоне увеличение его численности.
Лесоводы, сообразуясь с экологическими свойствами популяций, разработали два вида рубок древостоя: промежуточного и главного пользования. Рубки промежуточного пользования проводятся во всех возрастах жизни леса с целью создания благоприятных условий для роста остающейся части древостоя. При рубках главного пользования убирается весь древостой, достигший возраста спелости.
Этологическая (поведенческая) структура - это система взаимоотношений между членами одной популяции. Этология (от греч. ethos - характер, нрав и logos - наука) - наука о биологических основах поведения животных, оформилась в 30-х годах XX века. Ее становление связывают с работами австрийского этолога Конрада Лоренца и нидерландского этолога Николаев Тинбергена, заложивших основы теории инстинктивного поведения. Усиление контактов этологии с популяционной экологией в 60-х годах XX века дало мощный толчок изучению социального поведения и этологической структуры популяций животных.
Поведение животных по отношению к другим членам популяции зависит от того, какой образ жизни ведут особи: одиночный или групповой. При одиночном образе жизни особи популяции независимы и обособлены друг от друга. Такой образ жизни характерен для многих видов, но лишь на определенных стадиях жизненного цикла. Полностью одиночное существование невозможно из-за основной жизненной функции - размножения. Хотя исключением могут быть отдельные водные животные с наружным способом оплодотворения, например, одиночные актинии. Виды с одиночным образом жизни иногда образуют временные скопления особей. Например, божьи коровки и жужелицы в сухой подстилке возле пней в местах зимовок, щуки и сомы на дне водоемов и т.д.
При групповом образе жизни животные образуют семьи, колонии, стаи и стада, которые организованы иерархически. Иерархия у животных представляет собой систему поведенческих связей между особями группы, регулирующую их взаимоотношения и доступ к пище, убежищу, особям противоположного пола. Иерархия и другие формы жизни группы носят приспособительный адаптивный характер и составляют сущность жизни вида.
Семья - простейшая группировка особей, которая после размножения может распадаться, а может состоять из родителей и потомков нескольких поколений, как, например, кланы у гиеновых собак, правды у львов. Различают семьи отцовского, материнского и смешанного типа, в зависимости от того, кто из родителей берет на себя уход за потомством. Например, самка темной саламандры обвивает кладку своим телом и не покидает ее до выклева личинок. Самец трехиглой колюшки после охраны кладки и заботы о ней еще некоторое время оберегает мальков. В семьях с устойчивым образованием пар, в охране и выкармливании принимают участие и самец, и самка. При семейном образе жизни территориальное поведение животных выражено наиболее ярко: различные сигналы, маркировки, ритуальные формы угрозы и прямая агрессия обеспечивают владение участком. Пение птиц, например, это разговорный язык, способ обозначения своей территории, привлечение внимания самки.
Колонии - групповое поселение оседлых животных. Они могут существовать длительно или возникать на период размножения как, например, у многих птиц: грачей, чаек, гагар, тупиков и других. Примером такого поселения может служить колония бакланов (Phalacrocorax bougainvillei), насчитывающая не менее 10 000 особей, в которой на 1 кв. м площади приходится не менее трех гнезд.
Стаи - временные подвижные группировки особей для защиты от врагов, добычи пищи, миграции. Наиболее широко стайность распространена среди саранчи, птиц и рыб. У млекопитающих она характерна для собачьих (волков, гиен, койотов и др.). У птиц стаи формируются при сезонных перелетах, у оседлых или кочевых форм - при зимних кормежках. В стаях между особями существует постоянная звуковая и зрительная связь. Например, пеликаны (Pelecanidae) интенсивнее добывают рыбу сообща, стаями загоняя ее на мелководье. Волчьи стаи образуются для групповой охоты зимой. При групповой охоте волки практикуют преследование с выходом на перехват жертвы, нагон жертвы на засаду или захват ее в кольцо. Для этого требуется согласованность и координация действий всех особей. В стаях млекопитающих велика роль вожаков.
Стада - это более длительные и постоянные объединения животных. Стадами, как правило, живут китообразные, копытные, приматы (например, павианы). Стадо представляет собой группу животных одного вида, сохраняющих близость друг к другу, ведущих себя одинаково. Размеры стада и его состав по возрасту и полу изменчивы во времени. Стада китов, обезьян включают десятки животных, стада северных оленей, сайгаков состоят из сотен и тысяч особей. Например, наиболее продуктивные стада северных оленей включают 300-400 особей, а для выживания африканских слонов в стаде должно быть не менее 25 особей.
В стадах осуществляются все основные функции жизни: добывание корма, защита от хищников, миграции, размножение, воспитание молодняка и т.п. Основу группового поведения животных в стадах составляют взаимоотношения доминирования-подчинения, основанные на индивидуальных различиях между особями. Среди доминантов в одних стадах выделяются лидеры - более опытные члены стада (как у северных оленей), в других стадах - вожаки, более сильные особи. Биологический смысл иерархической системы доминирования-подчинения заключается в создании согласованного поведения группы, выгодного всем ее членам.
Эффект группы. Группам присущи все типичные черты популяций, но они характеризуются более высокой степенью объединения и способностью к саморегуляции численности. Это свойство носит название эффекта группы. Одним из показателей эффекта группы является территориальность. Многие животные привязаны к определенным местам. Они метят границы своей территории, на которой добывают пищу, строят гнезда или жилище, выкармливают молодняк. Положительное значение территориальности заключается в том, что она предупреждает перенаселенность.
У животных, ведущих групповой образ жизни, наблюдается тесное общение особей посредством запахов, звуков, специфики поведения. Благодаря сложной системе сигнализации у особей и их взаимному обмену информацией возрастает эффективность функционирования группы, направленная на удовлетворение важных жизненных потребностей всех ее членов.
Положительный эффект группы проявляется лишь до некоторого оптимального уровня плотности популяции.
Если особей становится слишком много, это оборачивается для них недостатком ресурсов среды. Тогда в действие вступают механизмы гомеостаза популяции.
В явлениях эффекта группы большую роль играют гормональные и нервные системы, регулирующие физиологические процессы в организме. Эффект группы проявляется как психофизиологическая реакция отдельной особи на присутствие других особей своего вида. Например, голуби некоторых пород не откладывают яйца, если не видят других птиц. Достаточно поставить перед самкой зеркало, чтобы она приступила к яйцекладке. У овец вне стада учащаются пульс и дыхание, а при виде приближающегося стада эти процессы нормализуются. Одиночно зимующие летучие мыши отличаются более высоким уровнем обмена веществ от живущих в колонии. Это ведет к повышенной трате энергии, истощению и часто заканчивается гибелью. Самое яркое проявление эффекта группы свойственно общественным насекомым. Пчелы, муравьи, термиты не могут жить изолированно, в одиночку. Они обладают сложной системой сигнализации, способствующей сохранению особей во времени и пространстве.
Таким образом, эффект группы проявляется у животных в ускорении темпов роста, в образовании условных рефлексов, повышении плодовитости, экономии энергетических затрат, повышенной средней продолжительности жизни индивидуума и т.д.
Лекция 8.
Динамика популяции
Динамика популяции - раздел популяционной экологии, изучающий численность особей популяции и механизмы ее регуляции. Жизнь популяции проявляется в ее динамике. К основным признакам динамики относятся:
- численность популяции - общее число особей на данной территории или в данном объеме (например, воды);
- плотность популяции - это среднее число особей на единицу площади или объема;
- рождаемость (плодовитость) - число новых особей, появившихся за единицу времени в результате размножения;
- смертность - число особей, погибших за определенный период;
- прирост популяции - разница между рождаемостью и смертностью;
- темп роста - средний прирост за единицу времени.
Изучение этих демографических признаков необходимо для выявления законов жизни популяции, а следовательно, и основ стабильности экосистемы в целом.
Численность популяции никогда не бывает постоянной и зависит от соотношения интенсивности размножения (плодовитости) и смертности. В процессе размножения происходит рост популяции, смертность же приводит к сокращению ее численности.
Рождаемость характеризует частоту появления новых особей в популяции. Различают рождаемость абсолютную и удельную, максимальную и экологическую.
Абсолютная рождаемость - число особей, появившихся в популяции за единицу времени. Удельная рождаемость выражается в числе родившихся особей на число особей в популяции в единицу времени. Например, для популяции человека показателем удельной рождаемости обычно служит число детей, родившихся в год на 1000 человек (таблица 4.2).
Максимальная рождаемость определяется числом самок в популяции и их способностью производить определенное число детенышей в единицу времени (т. е. физиологической плодовитостью). Обычно рождаемость ниже максимальной, т.к. она соответствует сложившимся экологическим условиям и называется экологической.
Таблица 4.2 Рождаемость и смертность в городе Смоленске
| Год | Абсолютная | Удельная | ||
| рождаемость | смертность | рождаемость | смертность | |
| 1991 | 3550 чел. | 3583 чел. | 10,7 чел. | 10,2 чел, |
| 1993 | 2644 чел. | 4480 чел. | 7,7 чел. | 12,7 чел. |
| 1994 | 2794 чел. | 4847 чел. | 7,9 чел. | 13,7 чел. |
Экологическая рождаемость дает представление о скорости размножения популяции при фактически сложившихся условиях жизни. Например, взрослая самка трески выметывает миллионы икринок, из которых в среднем доживают до взрослого состояния лишь 2 особи. В результате гибели по разным причинам (болезни, паразиты, хищники и др.) яиц, гусениц и куколок на свет появляется лишь 0,32% взрослых бабочек от числа отложенных яиц.
Численность и плотность популяции зависят также от ее смертности. Смертность популяции - это количество особей, погибших за определенный период. Она, как и плодовитость, изменяется в зависимости от условий среды, возраста и состояния популяции и выражается в процентах к начальной или, чаще, к средней величине ее. У большинства видов смертность в раннем возрасте всегда выше, чем у взрослых особей. У многих рыб до взрослой фазы доживает 1-2% от выметанной икры; у насекомых - 0,3-0,5% от отложенных яиц. Смертность, как и рождаемость, может быть абсолютной и удельной (таблица 4.2).
Выживаемость - средняя для популяции вероятность сохранения особей каждого поколения за определенный промежуток времени. Различают три типа смертности или, как их чаще называют, «кривые выживаемости». Каждый вид имеет свою кривую выживаемости (рис. 11).
Первая кривая - сильно выпуклая. Выпуклость кривой характеризует повышение смертности к концу жизни, остававшуюся до этого низкой. Такой тип кривой характерен для насекомых, личинки которых обитают в почве, воде, древесине или других местах с благоприятными условиями. Он характерен также для проходных рыб, нерестящихся один раз в жизни, для многих видов крупных животных и для человека
Вторая кривая - сильно вогнутая. Вогнутость кривой характерна для видов, смертность у которых очень высока на ранних стадиях жизни. Этот тип кривой свойственен большинству растений и животных. Максимальная гибель многих растений происходит в стадии прорастания семян или всходов, а животных - в личиночной фазе или молодом возрасте, например, у устриц, рыб, птиц, многих беспозвоночных.
Третий тип кривой - промежуточный, почти прямая линия, характерна для видов, у которых смертность мало изменяется с возрастом и остается более или менее одинаковой в течение всей жизни данной группы. Такая смертность встречается очень редко и только у популяций, постоянно находящихся в оптимальных условиях, например, таких, как гидра пресноводная.

Рис. 11. Три типа кривых выживания:
1 - человек, 2 - рыбы, 3 - гидра
Форма кривой выживания связана со степенью заботы о потомстве и способами защиты молоди. Так, кривые выживания пчел и дроздов, которые заботятся о потомстве, менее вогнуты, чем кривые выживания кузнечиков или сардин, не заботящихся о потомстве.
В замкнутых популяциях (в которых нет миграций) скорость изменения численности определяется только соотношением рождаемости и смертности. Если рождаемость выше смертности, то удельная скорость роста положительная. Если же смертность выше рождаемости, то удельная скорость становится отрицательной и численность популяции начинает убывать. Рождаемость и смертность, т. е. динамика численности, напрямую связаны с возрастной и половой структурами популяции.
Популяция регулирует свою численность и приспосабливается к изменяющимся условиям среды путем обновления и замещения особей. Особи появляются в популяции благодаря рождению и миграции, а исчезают в результате смерти и эмиграции.
При сбалансированной интенсивности рождаемости и смертности формируется стабильная популяция, в которой смертность компенсируется приростом и численность ее, а также ареал поддерживаются на одном уровне.
Популяции, в которых рождаемость превышает смертность и численность популяции растет так быстро, что наступает вспышка массового размножения, называются растущими. Это особенно характерно для мелких животных. Примером может служить увеличение численности популяции колорадского жука (Leptinotarsa decemlineata), быстро расселившегося на территории от Франции до Украины, Белоруссии, Смоленской и Псковской областей. Примером растущей популяции является элодея, завезенная из Америки, появившаяся в 1836 году в Ирландии, и проникшая уже в 1885 году в бассейн Оки. В последние десятилетия наметился рост популяции канареечного вьюрка (Serinus canaria), зеленой пеночки (Phylloscopus trochiloides), чайки обыкновенной (Larus ridibundus), зайца-русака и других видов.
Однако при бурном развитии популяции наступает переуплотнение, что ведет к ухудшению условий существования. А это приводит к резкому возрастанию смертности, в результате чего численность популяции начинает сокращаться. Если смертность превышает рождаемость, популяция становится сокращающейся. Так произошло, например, с элодеей и домовым воробьем (Passer domesticus) в умеренной зоне. В сокращении численности популяций многих животных часто повинен человек; например, таких, как соболь (Martes zibellina), бобр (Castor fiber), зубр (Bison bonasus), дрофа (Otis tarba) и других. Однако сокращаться безгранично популяция также не может. При определенном уровне численности интенсивность смертности начинает падать, а плодовитость повышается. В итоге сокращающаяся популяция превращается в растущую.
В природе численность популяций всегда испытывает колебания. Амплитуда и период этих колебаний зависят от вида и от условий среды обитания. Различают непериодические (нерегулярные, хаотические) и периодические (регулярные, циклические) колебания численности популяций. К непериодическим колебаниям численности, а соответственно и плотности популяции, относятся вспышки массового размножения непарного шелкопряда (Ocneria dispar) в южной и юго-восточной частях России в 1879 году, рыжего соснового пилильщика (Neodiprion sertifer) в Ленинградской и Смоленской областях, в Белоруссии с 1958 по 1962 годы. Резкий подъем численности наблюдается у популяций, оказавшихся на новом местообитании. Например, массовое размножение кроликов и разрастание зарослей кактуса опунции в Австралии, колорадского картофельного жука (Leptinotarsa decemlineata) и чайки обыкновенной (Larus ridibundus) в последние годы в Смоленской области.
Периодические колебания повторяются через равные промежутки времени, обычно в течение нескольких лет или одного сезона. Например, циклические изменения с подъемом численности в среднем через 4 года зарегистрированы у леммингов (Dicrostohyx), полярной совы (Nyctea scandiaca) и других животных тундры. Сезонные колебания численности характерны для многих насекомых, мышевидных грызунов, птиц. На периодические колебания численности популяции одним из первых обратил внимание русский генетик С.С. Четвериков (1880—1959), исследовавший изменчивость в природных популяциях. Колебания численности особей, составляющих популяцию, получили название популяционных волн (рис. 12).

Рис. 12. Популяционные волны
Численность и плотность - основные параметры, которые выражают количественные характеристики популяции как целого.
Каждой популяции свойственен так называемый биотический потенциал - способность к увеличению численности за данный промежуток времени. У разных организмов биотический потенциал не одинаков. Примером организмов с высоким потенциалом размножения являются:
- бактерии Bacillus coli, размножающиеся простым делением каждые 20 минут, которые при наличии условий для реализации биотического потенциала, могли бы освоить все пространство земного шара за 36 часов;
- гриб-дождевик, приносящий до 7,5 млрд спор, уже во втором поколении освоил бы всю Землю.
Крупным организмам с низким потенциалом размножения потребовалось бы для этого несколько десятилетий или столетий.
Таким образом, численность каждого вида при благоприятных условиях способна расти по так называемой экспоненциальной (логарифмической) кривой. Рост численности в геометрической прогрессии называется экспоненциальным ростом. График зависимости численности популяции от времени при экспоненциальном росте представляет собой кривую, напоминающую по форме латинскую букву J, называемую зкспонентой. Эта J-образная кривая показывает, что в ходе роста популяции ее численность увеличивается с возрастающей скоростью (рис. 13).

Рис. 13. Экспоненциальная кривая роста популяции
Величина изменения численности за единицу времени называется абсолютной скоростью роста численности. Эта величина зависит от численности популяции: например, из графика на рис. 14 видно, что чем больше численность, тем больше абсолютная скорость роста. Удельная скорость роста численности - это скорость прироста на единицу особи - эта величина и отражает биотический потенциал.
Экспоненциальный рост в реальных условиях наблюдается, когда популяция растет в условиях избытка ресурсов (пищи, места для размножения), отсутствия конкурентов и не испытывает воздействия неблагоприятных факторов. В лабораторных условиях экспоненциальный рост можно наблюдать у популяций микроорганизмов (дрожжей, бактерий, хлореллы) в начальной фазе их роста. В природе экспоненциальный рост наблюдается при вспышке численности грызунов, саранчи, непарного шелкопряда и других насекомых. Экспоненциально может расти численность популяций, вселенных в новую местность, где у них много нищи и мало врагов. Классическим примером такого роста является рост численности кроликов, завезенных в Австралию. Примером экспоненциального роста можно считать размножение микроорганизмов в загрязненных органическими и биогенными веществами водоемах.

Рис. 14. Рост населения мира (млн чел.)
Близок к экспоненциальному типу рост населения популяции человека в настоящее время (рис. 15). Он обусловлен прежде всего резким снижением смертности в детском возрасте.

Рис. 15. Логистическая кривая роста популяции
Естественный рост популяции никогда не реализуется в форме экспоненциальной кривой. В крайнем случае, если это и происходит, то в течение относительно короткого отрезка времени (как в приведенных выше примерах), после чего скорость роста численности снижается. Объясняется это тем, что не только в природных, но и в оптимальных экспериментальных условиях рост численности ограничен комплексом факторов внешней среды и реально складывается как результат соотношения меняющихся значений рождаемости и смертности.
Рост численности популяции замедляется по мере увеличения ее плотности, так как условия для роста и размножения особей становятся менее благоприятными. Например, животным при высокой плотности популяции может не хватать пищи. Растения начинают затенять друг друга, или им не хватает влаги. По мере ухудшения условий удельная скорость роста снижается, и при некоторой плотности численность популяции перестает расти. Эту предельную плотность, которой может достигнуть популяция в данных условиях, называют емкостью среды. Если рост популяции ограничен ресурсами, то после достижения емкости среды численность популяции колеблется возле некоторого среднего уровня и популяция подчиняется правилам логистического роста.
График изменения численности популяции при логистическом росте представляет собой кривую, которая называется логистической кривой и напоминает по форме латинскую букву S (рис. 15). При логистическом росте популяции ее численность некоторое время нарастает, но вскоре этот процесс начинает замедляться, и постепенно рост численности практически прекращается. Для большинства популяций и видов выживаемость отображается логистической кривой.
Лекция 9.
Гомеостаз популяции
Популяции организмов обладают способностью к естественному регулированию плотности. Плотность популяции при более или менее значительных колебаниях остается в устойчивом состоянии между своими верхним и нижним пределами. Это обеспечивается действием определенных приспособительных механизмов.

Рис. 16. Равновесное состояние популяции, определяемое соотношением факторов, увеличивающих или снижающих ее численность
Тенденция популяций поддерживать внутреннюю стабильность с помощью собственных регулирующих механизмов называется гомеостазом, а колебания численности популяций в пределах какой-то средней величины - их динамическим равновесием. Все биологические системы характеризуются способностью к гомеостазу, т.е. к саморегуляции. С помощью саморегуляции поддерживается в целом существование каждой системы - ее состав и структура, внутренние связи и преобразования в пространстве и во времени.
Гомеостатическими являются прежде всего отдельные особи, а затем популяции. Саморегулирующиеся системы не замкнуты, они активно взаимодействуют с внешней средой и, следовательно, подвержены изменениям. Саморегуляция - необходимое приспособление организмов для поддержания жизни в постоянно меняющихся условиях. Саморегулирование популяции осуществляется действующими в природе двумя взаимно уравновешивающимися буферными силами. С одной стороны, это биотический потенциал, составляющий совокупность всех факторов, способствующих увеличению численности популяции, а с другой - это сопротивление среды - совокупность факторов, снижающих численность популяции (рис. 16).
Итак, рост, снижение или постоянство численности популяции зависят от соотношения между биотическим потенциалом (прибавлением особей) и сопротивлением среды (гибелью особей). Изменения численности популяции какого-либо вида - это результат нарушения равновесия между ее биотическим потенциалом и сопротивлением окружающей ее среды. Равновесие - понятие относительное. Например, в какой-то год популяция может снизить свою численность из-за засухи, а в последующие годы с нормальным увлажнением полностью восстановить ее. Подобные циклические колебания обычно продолжаются неопределенно долго.
Биологическое регулирование популяции (или ее автоматическое саморегулирование), абиотические факторы, независимые от плотности популяции, вызвать не могут, если они действуют изолированно от биотических. Зависимая от плотности динамика популяций обеспечивается биотическими факторами. Их называют регулирующими. Они «работают» по принципу обратной отрицательной связи: чем значительнее численность, тем сильнее срабатывают механизмы, обусловливающие ее снижение, и наоборот, - при низкой численности сила этих механизмов ослабевает и создаются условия для более полной реализации биотического потенциала. Факторы такого типа лежат в основе популяционного гомеостаза, обеспечивающего поддержание численности в определенных границах значения.
Примером регулирующих факторов являются взаимоотношения организмов типа хищник - жертва, паразит - хозяин и конкуренция. Высокая численность жертвы создает условия (пищевые) для размножения хищника. Последний, в свою очередь, увеличив численность, снижает количество жертвы. Численность обоих видов в результате этого носит синхронно-колебательный характер.
Действие регулирующих факторов можно рассматривать на уровне межвидовых и внутривидовых (внутрипопуляционных) взаимоотношений организмов. К межвидовым механизмам гомеостаза относятся взаимоотношения хищник - жертва, паразит - хозяин и конкуренция. Конкуренция лежит и в основе внутрипопуляционного гомеостаза. Она может проявляться в жестких и смягчающих формах. Жесткие формы заканчиваются гибелью части особей.
Например, в растительном мире внутривидовая конкуренция проявляется в самоизреживании фитоценозов. На стадии всходов и молодых растений в лесных сообществах на 1 га насчитывается до нескольких сотен тысяч древесных растений. К возрасту спелости (100-120 лет для хвойных видов и 50-70 лет для лиственных) число экземпляров обычно не превышает 1000 на 1 га, но чаще - несколько сотен. Остальные погибают в результате острой конкуренции (рис. 17).

Рис. 17. Ход самоизреживания леса с возрастом
В животном мире результат острой внутривидовой борьбы проявляется в форме каннибализма (поедания себе подобных). Например, явления каннибализма характерны для некоторых грызунов, личинок насекомых (майского жука), хищных рыб - щук, окуней, корюшки, трески, наваги и других.
Смягченные формы внутривидовой конкуренции проявляются обычно через ослабление части особей, выключение их из процессов размножения.
К механизмам внутрипопуляционного гомеостаза относятся угнетающие (ингибирующие) выделения во внешнюю среду более сильными особями, стрессовые явления, территориальность, миграции между популяциями. Выделения во внешнюю среду характерны как для растительных, так и для животных организмов. На лабораторных животных (крысы, мыши) показано, что перекачивание воздуха из перенаселенных помещений в те помещения, где животные свободно размножаются и нормально развиваются, приводит к замедлению роста и угнетению этих животных. Аналогичные результаты наблюдались в опытах с головастиками лягушек, когда в аквариум, где они хорошо развивались, добавляли воду из аквариумов, в которых плотность организмов была чрезвычайно высокой.
Явления территориальности наиболее четко выражены в животном мире. К ним относятся различные способы охраны занимаемых территорий. У кошачьих и собачьих территориальность проявляется через мечение границ участков выделением желез, мочой или механическими отметинами на деревьях и почве. Пение птиц - это прежде всего сигнал о занятости территории в период размножения и последующего выкармливания потомства.
Важным механизмом регуляции численности, проявляющимся в переуплотненной популяции, является стресс-реакция (от англ. stress - напряжение). Если на популяцию воздействует какой-то сильный раздражитель, она отвечает на него неспецифической реакцией, называемой стрессом. В живой природе различают много форм стресса: антропический (возникает у животных под воздействием деятельности человека), нервно-психический (проявляется при несовместимости индивидуумов в группе или в результате переуплотнения популяции), шумовой и другие.
Например, в результате переуплотнения популяции, у отдельных особей возникают физиологические изменения, которые приводят к резкому сокращению рождаемости и увеличению смертности. У млекопитающих такое явление получило название стресс-синдрома. При этом животные становятся настолько агрессивными (жестокие драки, нетерпимость присутствия соседа и т.д.), что у них почти полностью прекращается размножение. У животных, находящихся в стрессовом состоянии, увеличивается кора надпочечников и повышается концентрация кортикостероидных гормонов. У самок нарушается овуляция, происходит резорбция эмбрионов, не проявляются инстинкты заботы о потомстве и т.д.
Миграции, как фактор гомеостаза могут проявляться в двух видах. Первый относится к массовому исходу особей из популяции при явлениях перенаселенности (нашествие). Такие явления характерны для леммингов, белок и других видов с взрывным типом динамики численности. Второй вид миграции связан с более постепенным (спокойным) уходом части особей в другие популяции с меньшей плотностью населения.
Гомеостаз в полной мере проявляется, если срабатывают все механизмы, лежащие в его основе. В настоящее время нарушения гомеостатических механизмов вызываются в большинстве случаев антропогенными факторами. В связи с этим, одной из важнейших задач человека является исключение или резкое снижение действия подобных факторов.
Лекция 10.
СИНЭКОЛОГИЯ- ЭКОЛОГИЯ СООБЩЕСТВ (БИОЦЕНОЗОВ)
Основные понятия синэкологии
В природе популяции разных видов интегрируются в макросистемы более высокого ранга - в сообщества, или биоценоаы. Биоценоз (от греч. bios - жизнь, koinos - общий) - это организованная группа популяций растений, животных, грибов и микроорганизмов, живущих совместно в одних и тех же условиях среды.
Понятие «биоценоз» было предложено в 1877 году немецким зоологом Карлом Мёбиусом. Изучая устричные банки Северного моря и приуроченные к ним организмы, Мёбиус пришел к выводу, что каждая отмель представляет собой сообщество живых существ, все члены которого находятся в тесной взаимосвязи. Биоценоз - не простая совокупность живых организмов, а согласованная, динамическая, уравновешенная, взаимосвязанная, стойкая во времени система, состоящая из фитоценоза (совокупность растений), зооценоза (совокупность животных), микоценоза (совокупность грибов) и микробоценоза (совокупность микроорганизмов) (рис. 18).

Рис. 18. Составные части биоценоза
Фитоценоз является экологическим каркасом биоценоза, обусловливающим его состав и структуру. Представляя собой ведущий структурный компонент биоценоза, фитоценоз определяет видовой состав зооценоза, микоценоза и микробоценоза.
Синоним биоценоза - сообщество. Выделяют три основных вида сообществ: наземные, пресноводные и морские. В природе существуют сообщества разного масштаба. Популяции разных видов сосуществуют в гниющем пне и в желудке коровы, на болоте и в лесу, на пространстве тайги или тундры.
Наиболее крупные сообщества, характеризующиеся определенным типом растительности и климатом, называются биомы (от англ. biome, греч. bios - жизнь и лат. oma - окончание, означающее совокупность), если речь идет об одном континенте, или типы биомов. Тайга, тундра, степи, саванна, пустыни, тропический дождевой лес - примеры типов биомов. Некоторые авторы биомы именуют формациями или комплексами. В представлении других биом - это преимущественно синтетическое понятие, характеризующееся почти устойчивым равновесием.
Сообщества, расположенные на участках с относительно однородной растительностью, называются биоценозы (например, биоценоз луга, биоценоз болота). По отношению к более мелким сообществам (население деревьев, муравейников, нор, разлагающихся пней и т.п.) применяют разнообразные термины: «микросообщества», «биоценотические группировки», «биоценотические комплексы», «синузии», «консорции» и другие. Биоценозы, созданные человеком, называются агроценозами (поле, сад, огород). Целостность биоценозов поддерживается эволюционно сложившейся системой связей, прежде всего информационных. Согласно концепции литовского эколога Э. Лекявичуса, на уровне биоценоза постоянно функционируют два канала информации. Один из них обеспечивает устойчивое существование и репродукцию популяций конкретных видов; это система самоподдержания и развития видов, или селфинг. Второй канал связывает биоценоз, как целое, с его компонентами; это координации, «принуждающие» популяции отдельных видов к выполнению специфических функций в составе целостной системы. На конфликте этих двух каналов информации строятся регуляторные механизмы, определяющие поддержание глобальных функций биоценоза.
Биоценозы, как биологические системы можно определить как исторически сложившиеся группировки живого населения биосферы, заселяющие общие места обитания, возникшие на основе биогенного круговорота и обеспечивающие его в конкретных природных условиях.
Как и популяция, сообщество имеет собственные свойства и показатели, присущие ему как целому. Свойствами сообщества являются устойчивость (способность противостоять внешним воздействиям) и продуктивность (способность производить живое вещество). Показателями сообщества являются характеристики его состава (разнообразие видов), структура пищевой сети, соотношение отдельных групп организмов. Одна из главных задач экологии - выяснить взаимосвязи между свойствами и составом сообщества, которые проявляются независимо от того, какие виды входят в него.
Структура биоценозов
Природное сообщество - биоценоз - имеет видовую, экологическую и пространственную структуры.
Видовая структура - один из важнейших признаков биоценоза. Ее главными показателями являются видовой состав - число видов и численность популяции - количественное соотношение особей.
Каждый конкретный биоценоз характеризуется определенным видовым составом. При этом одни виды биоценоза могут быть представлены многочисленными популяциями, а другие - малочисленными. Число видов в составе биоценоза, приходящихся на единицу площади, называется его видовой насыщенностью. Виды, преобладающие в биоценозе, называются доминантами (от лат. dominantie - господствующий). Например, в еловых лесах среди деревьев доминирует ель, в травяном покрове - кислица или зеленые мхи, а среди птиц - королек или зарянка.
Наряду с доминантами в биоценозах выделяют виды-эдификаторы (от лат. aedificator - строитель). Эдификаторы - это строители биоценоза, т. е. виды, сильно изменяющие среду и тем создающие определенные условия для жизни других видов данного биоценоза. Обычно вид-доминант одновременно является и эдификатором. Например, ель в еловом лесу наряду с доминантностью обладает высокими эдификаторными свойствами. Это выражается в ее способности сильно затенять почву, создавать кислую среду своими корневыми выделениями и образовывать подзолистые почвы. Вследствие высоких эдификаторных свойств ели под ее пологом могут жить только теневыносливые и тенелюбивые виды растений. Под пологом елового леса доминантой может быть черника, но она не является эдификатором. В сосновых лесах эдификатором является сосна. Но, по сравнению с елью, она более слабый эдификатор, т. к. сосновый лес относительно светлый и редкоствольный. На сфагновых торфяниках эдификаторами являются сфагновые мхи, в дубраве - дуб, в ковыльной степи - ковыль и т.д.
По общему числу видов и их соотношению судят о видовом разнообразии биоценоза. Видовое разнообразие - признак экологического разнообразия: чем больше видов, тем больше экологических ниш в данном сообществе.
Экологическая структура биоценоза - это соотношение экологических групп организмов, выполняющих в сообществе в каждой экологической нише определенные функции. Каждый биоценоз слагается из определенных экологических групп организмов. Например, в сухих аридных условиях доминируют склерофиты и суккуленты, а на увлажненных территориях - гигрофиты.
Экологическую структуру биоценоза отражает также соотношение групп организмов, объединяемых сходным типом питания. Например, в лесах преобладают сапрофаги, в степях - фитофаги, в глубинах Мирового океана - хищники и детритофаги. Биоценозы со сходной экологической структурой могут иметь разный видовой состав, так как в них одни и те же экологические ниши могут быть заняты сходными по экологии, но не родственными видами. Такие виды выполняют одни и те же функции в сходных биоценозах и называются викарирующими или замещающими. К примеру, викарирующими видами являются сайгаки в степях Казахстана, антилопы в саваннах Африки, бизоны в прериях Америки, кенгуру в саваннах Австралии. Они занимают сходные экологические ниши и выполняют одни и те же функции.
Пространственная структура выражается в горизонтальном и вертикальном расчленении фитоценоза на отдельные элементы, каждый из которых играет свою роль в накоплении и преобразовании вещества и энергии. По вертикали растительное сообщество разделяется на ярусы - горизонтальные слои, толщи, в которых располагаются надземные и подземные части растений определенных жизненных форм. Ярусность особенно четко выражена в лесных фитоценозах, где выделяют до шести ярусов:
I ярус - деревья первой величины (ель, сосна, береза, липа, клен,дуб);
II ярус - деревья второй величины (рябина, черемуха, ива);
III ярус - кустарники (лещина, шиповник, бересклет, жимолость);
IV ярус - кустарнички и высокие травы (багульник, вереск, голубика, иван-чай);
V ярус - мелкие травы (кислица, осока, копытень европейский и др.);
VI ярус - мхи, напочвенные лишайники, печеночники.
Малоярусные сообщества (луг, степь, болото) имеют по два-три яруса. Ярусно располагаются и подземные части растений. Корни у деревьев проникают глубже, чем у кустарников, ближе к поверхности располагаются корни травянистых растений, а непосредственно на ней - ризоиды мхов. При этом в поверхностных слоях почвы корней значительно больше, чем в глубинных.
Благодаря ярусности различные растения, особенно их органы питания (листья), располагаются на разной высоте и поэтому легко уживаются в сообществе. Таким образом, ярусы определяют сложение и структуру фитоценоза. Если их мало, то растительное сообщество называют простым, если много - сложным.
Растения каждого яруса и обусловленный ими микроклимат создают определенную среду для животных и микроорганизмов. Например, в почвенном ярусе леса обитают бактерии, грибы, насекомые, клещи, черви. Более высокие ярусы занимают растительноядные насекомые, птицы, млекопитающие. При этом птицы обитают в строго определенном ярусе, особенно в период гнездования. Так, к видам, гнездящимся только на земле, относятся куриные, овсянки, тетерева. В кустарниковом ярусе обитают певчие дрозды, славки, снегири, а в кронах деревьев - зяблики, корольки, щеглы, хищные птицы и другие.
Животное население биоценоза (зооценоз), будучи приурочено к растениям, также распределяется по ярусам. Например, среди насекомых выделяют следующие группы:
- геобий - обитатели почвы;
- герпетобий - насекомые наземного поверхностного слоя;
- бриобий - обитатели мохового яруса;
- филлобий - обитатели травостоя;
- аэробий - обитатели более высоких ярусов.
В водных сообществах вертикальная ярусная структура в первую очередь обусловлена внешними условиями, а именно световым и температурным режимами. Например, в водном сообществе выделяют следующие ярусы:
- полупогруженные растения;
- укореняющиеся растения с плавающими листьями;
- подводные растения;
- придонные растения.
Среди организмов имеются и внеярусные, например, лианы, различные эпифиты, паразиты, многие животные, свободно переходящие из одного яруса в другой. Особенно много таких организмов во влажных тропических лесах, структура которых очень сложна.
Виды животных и растений разных ярусов различают по их отношению к условиям среды. Так, растения каждого нижележащего яруса более теневыносливы, чем расположенные над ними. Виды различных ярусов в биоценозе находятся в тесных взаимоотношениях и взаимозависимости. Растения нижних ярусов положительно влияют на животное население как в количественном, так и в качественном соотношениях.
Ярус можно рассматривать как структурную единицу биоценоза, отличающуюся от других частей его определенными экологическими условиями, набором растений, животных и микроорганизмов. В каждом ярусе складывается своя система взаимоотношений составляющих его компонентов.
Вертикальное распределение организмов в биоценозе обусловливает и определенную структуру в горизонтальном направлении.
Горизонтальная структура биоценозов выражена их мозаичностью и реализуется в виде неравномерного распределения популяций отдельных видов по площади. Это определяется, с одной стороны, неодинаковостью почвенно-грунтовых условий и микроклимата, а с другой - взаимоотношениями отдельных особей как внутри популяции, так и между собой. На этой основе формируются разного рода микрогруппировки, в которых видовые популяции связаны между собой более тесными функциональными отношениями, чем с остальной частью биоценоза.
К таким микрогруппировкам относятся консорции, синузии, парцеллы. Консорция (от лат. consortium - соучастие) - функциональная единица биоценоза, включающая самостоятельно существующее зеленое растение и все организмы, связанные с ним трофическими, топическими, форическими и фабрическими отношениями. Самостоятельно существующее автотрофное растение является центром консорции, или детерминантой, по отношению к которой связанные с ней виды именуются консортами. Консортами могут быть паразиты-растения, грибы, эпифиты, лианы, птицы, насекомые, животные-фитофаги. Например, дерево липа является центром консорции или детерминантой, а ее консортами могут быть: личинки хруща - потребители корней; микориза на корнях; жук-короед; жук-листоед; гусеница шелкопряда, питающаяся листвой дерева; пчела - опылитель цветков; гнездо дрозда; лось - потребитель веточного корма; лесная мышь - потребитель семян и другие. Среди консорций выделяют несколько типов: индивидуальные (одного растения), ценопопуляционные (популяции вида в растительном сообществе), региональные и видовые.
Синузии (от греч. synusia - совместное пребывание, сообщество) - пространственно и экологически разграниченные части фитоценоза, состоящие из видов растений одной или нескольких экологически близких жизненных форм. Если ярус - морфологическое понятие, то синузия - экологическое. Она может совпадать с ярусом и может составлять только часть его. В ярусе может быть несколько синузий. В синузиях отражена мозаика экологических факторов формирования растительного сообщества: сосна занимает сухие песчаные почвы, ель - более влажные супесчаные и суглинистые, береза и осина - вырубки и т.д. Например, в полынно-солянковой пустыне можно выделить синузии ранневесенних эфемеров и синузии летне-осенних полукустарников (полыни, солянки); в сосновом лесу - синузии брусники и черники.
Парцеллы - это структурные части горизонтального расчленения биоценоза, отличающиеся составом, структурой, свойствами компонентов, спецификой их связей и материально-энергетического обмена. Парацеллярные, т. е. элементарные, группировки выделяются не только в популяциях растений, но и в популяциях животных в отличие от синузии и яруса, рассматриваемых обычно как понятия ботаники.
Пространственная структура сообщества является показателем имеющегося в данном местообитании разнообразия экологических ниш, богатства и полноты использования сообществом ресурсов среды, а также показателем относительной устойчивости сообщества.
Лекция 11.
Отношения организмов в биоценозах
Основу возникновения и существования биоценозов представляют взаимоотношения организмов и их разнообразные связи, которые невероятно сложны. В первую очередь это внутривидовые связи, межвидовые отношения всех типов и трофические связи. В биоценозе организмы связаны также топическими, форическими и фабрическими отношениями, которые выделил русский зоолог В.Н. Беклемишев.
Отношения между организмами в биоценозе определяются также временем их нахождения в сообществе. Они могут быть постоянными (сессильными) и временными (вагильными). Постоянство характерно для растений, животные чаще пребывают в сообществе временно - в течение суток, сезона или в период миграции. Ученые подсчитали, что если в сообществе 1000 видов, то число связей между ними теоретически может доходить до 499 500.
Трофические (пищевые) связи среди всех взаимоотношений между организмами имеют первостепенное значение. Любой биоценоз включает несколько трофических (пищевых) уровней, или звеньев, которые образуют трофическую структуру сообщества (рис. 19).

Рис. 19. Модель трофической структуры сообщества
Первый уровень представлен растениями, которые называются автотрофами или продуцентами (от лат. producentis - создающий). Второй уровень представлен животными, называющимися гетеротрофами, - фитофагами или консументами первого порядка. Третий уровень (иногда четвертый, пятый) представлен хищниками (зоофагами), или консументами второго (третьего, четвертого) порядка. Последний уровень в основном представлен микроорганизмами и грибами, питающимися мертвым веществом. Их называют сапрофагами или редуцентами (от лат. reducens - возвращать).
Взаимосвязанный ряд трофических уровней представляет цепь питания, или трофическую цепь. Любое сообщество можно представить в виде пищевой сети, т.е. совокупности трофических взаимосвязей между видами этого сообщества. Пищевая сеть (ее переплетения бывают очень сложными) обычно состоит из нескольких пищевых цепей, каждая из которых является отдельным каналом, по которому передаются и вещество, и энергия (рис. 20).
В каждом звене большая часть энергии расходуется в виде тепла, теряется, что ограничивает число звеньев в цепи.
Большинство цепей начинается растением, а заканчивается хищником, причем наиболее крупным.
Обычно цепь питания состоит из трех-четырех-пяти звеньев. Например:
растение - заяц - волк;
растение - полевка - канюк;
растение - гусеница - синица - ястреб;
растение - насекомое - лягушка - гадюка - орел.
Почти все животные, за исключением узкоспециализированных видов, используют разнообразные источники пищи. Чем больше видовое разнообразие и богаче пищевые связи, тем устойчивее биоценоз.
В биоценозах различают два типа трофических сетей: пастбищную и детритную.

Рис. 20. Детритная пищевая цепь (схема)
В пастбищной сети, или сети выедания, поток энергии идет от растений к растительноядным животным, а далее к консументам более высокого порядка.
В детритной сети, или сети разложения, поток энергии начинается с мертвых растительных и животных остатков, экскрементов и идет к первичным детритофагам - редуцентам, разлагающим органические вещества (рис. 20).
Трофические цепи и сети составляют основу взаимосвязей в живой природе, но пищевые связи - это не единственный вид взаимоотношений между организмами. В системе биоценотических отношений выделяются топические связи, вытекающие из пространственной структуры. К этой категории взаимоотношений относится борьба за место для поселения, конкуренция животных за убежища. Эти связи крайне разнообразны.
При борьбе за место поселения может происходить либо расширение круга пригодных для заселения биотопов (эвритопность), либо высокая степень специализации (стенотопность), что уменьшает число конкурентов. Примером стенотопности могут служить пищухи Certhia familiaris - птицы, строящие гнезда в узких клинообразных щелях стволов деревьев и поэтому практически не имеющие конкурентов по части выбора мест для гнездования.
Большая роль в создании или изменении среды для других организмов принадлежит растениям. Они участвуют в формировании мезо- и микроклимата, влияя на температурный режим, освещенность, влажность. Растения создают вокруг себя биохимическую среду, влияющую на другие виды, растущие по соседству. Например, заросли тростника и водной растительности, снижая течение и силу ветра, создают условия для поселения ряда видов животных, для которых существование на открытых водоемах невозможно.
В водных биоценозах скопление водорослей создают как условия для поселения многих видов беспозвоночных животных, так и убежище для разных видов рыб. Животные строят норы, гнезда и другие сооружения, создают убежища с благоприятным микроклиматом, используемые и другими видами. Нора млекопитающих привлекает целый ряд видов членистоногих, амфибий и других животных, использующих их вместе с хозяевами.
Строительная деятельность животных может иметь и более широкий биоценотический эффект. Например, плотины бобров изменяют гидрологические свойства лесных рек и ручьев, что ведет к изменению растительности в зоне бобровых прудов, образованию аллювиальных кос низке плотины, заилению затопленных участков леса и т.д. Бобровые пруды создают особые условия и для жизни животных; формируется комплекс водных и околоводных сообществ, не свойственных исходному типу биоценоза.
В.Н. Беклемишев в системе биоценотических отношений выделил также форические связи (от лат. foras - вне, за пределами), выражающиеся в расселении (распространении) одного вида другими.
В роли транспортировщиков выступают животные. Перенос животными плодов, семян, пыльцы растений называется зоохорией (от греч. zoon - животное, choreo - двигаюсь, распространяюсь). Различают эндозоохорию и эктозоохорию. Эндозоохория - это распространение семян через пищеварительный тракт животных. У большинства птиц и многих млекопитающих семена, проходя через пищеварительный тракт, сохраняют жизнеспособность и таким образом распространяются на далекие расстояния. Например, птицы таким способом распространяют семена рябины, облепихи. Воробьи разносят семена земляники, крушины, жимолости и других ягодников. Большую роль в распространении растений играют перелетные птицы. Благодаря им растения северных областей попадают в степи. С деятельностью птиц связывают появление большинства деревьев на океанических островах. Овцы, лошади и другие домашние животные разносят семена травянистых растении, такие как пастушья сумка, овсюг, дикая редька, мокрица.
Активная эктозоохория связана с запасанием животными корма и переносом его на значительные расстояния. Например, кедровка (Nucifraga caryocatactes) разносит семена кедровой сосны (Pinus sibirica). Большие запасы кедровых орешков делает бурундук (Eutamias sibiricus). Однако не все семена используются в пищу - либо из-за гибели зверьков, либо из-за потери. Это способствует прорастанию уцелевших семян в местах, удаленных от растения, которому они принадлежали. В расселении дуба (Quercus robur) участвуют сойки (Carrulus glandarius). В распространении лесного орешника лещины (Corylus avellana) участвуют белки (Sciurus vulgaris). Они растаскивают орехи по всему лесу. В результате лещина расселяется на огромных площадях.
Мелкие мышевидные грызуны также запасают много семян на зиму. Например, лесная мышь (Apodemus sylvaticus) за шесть дней может утащить до 1500 желудей. Распространению ряда растений способствуют муравьи. Подсчитано, что одна колония муравьев (Formica rufa) рассеивает на расстоянии до 70 метров от муравейника свыше 36 000 семян различных растений.
Пассивная эктозоохория заключается в том, что семена растений транспортируются на значительные расстояния, прикрепляясь с помощью специальных приспособлений к телу животных.
К форическим связям, кроме зоохории, относится и форезия (от лат. foras - за пределами) - перенос животными других более мелких животных. Такие связи существуют, например, между крупными позвоночными и поселяющимися на их теле организмами-обрастателями (гидроидные полипы, некоторые моллюски, водоросли и другие). Малярийные комары являются переносчиками возбудителей малярии; млекопитающие переносят клещей, блох и других паразитов.
Фабрические связи (от лат. fabrico - изготовлять) - тип отношений в биоценозе, при которых организм использует для создания своих сооружений мертвые остатки, продукты выделения либо даже части и органы других организмов. Например, птицы при строительстве гнезд используют сухие травинки и веточки, шерсть и пух млекопитающих; рак-отшельник - раковины мертвых брюхоногих моллюсков в качестве убежища. Бобр создает плотины и хатки из растительного материала. Личинки ручейников строят домики из кусочков веток, коры растений, из раковин мелких видов катушек, захватывая далее раковинки с живыми моллюсками.
Лекция 12.
Типы биотических отношений
Биотические факторы - это совокупность влияния жизнедеятельности одних организмов на другие. Эти факторы очень разнообразны и проявляются во взаимоотношениях организмов при совместном обитании. Среди огромного многообразия взаимосвязей организмов можно выделить определенные типы отношений, имеющие много общего у организмов самых разных систематических групп. Основными типами биотических отношений являются: конкуренция, хищничество, комменсализм, мутуализм, синойкия, паразитизм.
Конкуренция (от лат. concurro - стучаться, сталкиваться) - взаимоотношения между организмами одного вида (внутривидовая конкуренция) или разных видов (межвидовая конкуренция), при которой они используют одни и те же ресурсы окружающей среды при их недостатке.
Чарльз Дарвин рассматривал внутривидовую конкуренцию как важнейшую форму борьбы за существование. Внутривидовая конкуренция повышает интенсивность естественного отбора. Примером внутривидовой конкуренции является самоизреживание у растений. У некоторых организмов (птиц, рыб и других животных) под влиянием внутривидовой конкуренции за пространство сформировался тип поведения, называемый территориальностью. Например, у птиц в начале сезона размножения самец выявляет участок обитания - территорию, которую он защищает от вторжения самцов того же вида.
Межвидовая конкуренция чаще всего проявляется между экологически близкими особями (или популяциями) разных видов. Она может быть пассивной (использование ресурсов окружающей среды, необходимых обоим видам) и активной (подавление одного вида другим). Классическим примером межвидовой конкуренции являются описанных русским биологом Г.Ф. Гаузе (1910-1986) опыты по содержанию популяций разных видов инфузорий-туфелек рода Paramecium на одном и том же ограниченном питании. Оказалось, что через некоторое время в живых остаются особи только одного вида, выжившие в борьбе за пищу, поскольку его популяция быстрее росла и размножалась. Вывод, сделанный Г.Ф. Гаузе на основании проведенных опытов, называют принципом конкурентного исключения Гаузе или правилом Гаузе. Оно гласит: «Два вида не могут устойчиво существовать вместе в ограниченном пространстве, если рост численности обоих ограничен одним жизненно важным ресурсом, количество и (или) доступность которого лимитированы». Другими словами, два вида не могут сосуществовать, если они занимают одну экологическую нишу.
Примером острой конкуренции может служить несовместимость в одном водоеме речных раков - широкопалого (Astacus astacus) и узкопалого (Astacus leptodactylus). Победителем оказывается узкопалый рак как наиболее плодовитый и приспособленный к современным условиям жизни. В Европе в поселениях человека серая крыса совершенно вытеснила другой вид того же рода - черную крысу, живущую в лесных и пустынных районах. Серая крыса крупнее, лучше плавает, агрессивнее, поэтому сумела победить. В России, наоборот, сравнительно мелкий рыжий таракан-прусак вытеснил более крупного черного таракана, потому что сумел лучше приспособиться к специфическим условиям человеческого жилья. В Австралии обыкновенная пчела, завезенная из Европы, вытесняет маленькую туземную, не имеющую жала.
У растений подавление конкурентов происходит в результате перехвата минеральных питательных веществ и почвенной влаги корневой системой, перехвата солнечного света листовым аппаратом, а также в результате выделения токсических соединений (аллелопатия). Основным ресурсом у растений является свет. Из двух сходных видов растений, сосуществующих в одной и той же среде, преимущества достигает тот вид, который способен раньше выйти в верхний, лучше освещенный ярус. Всходы ели хорошо развиваются под защитой сосен, берез, осин, но впоследствии, при разрастании еловых крон, всходы этих светолюбивых пород гибнут. Сорняки угнетают культурные растения посредством перехвата влаги и минеральных питательных веществ, затенения и аллелопатии.
Победителем в конкурентной борьбе оказывается тот вид, который в данной экологической обстановке имеет хотя бы небольшие преимущества перед другими, т. е. больше приспособлен к условиям окружающей среды. Причины вытеснения одного вида другим могут быть различны, но поскольку экологические спектры даже близких видов никогда не совпадают полностью, даже при общем сходстве требований к среде, виды все же чем-либо отличаются друг от друга. В результате конкуренции в сообществе совместно уживаются только те виды, которые сумели хотя бы немного разойтись в экологических требованиях. Так, насекомоядные птицы, кормящиеся на деревьях, избегают конкуренции друг с другом благодаря разному характеру поиска добычи на разных частях дерева. В африканских саваннах копытные используют пастбищные корма по-разному: зебры обрывают верхушки трав, антилопы гну поедают определенные виды растений, газели выщипывают только низкие травы, а антилопы топи кормятся высокими стеблями.
Таким образом, межвидовая конкуренция может иметь два итога: либо вытеснение одного из двух видов из сообщества, либо расхождение обоих видов по экологическим нишам. Конкуренция - один из важнейших факторов формирования видового состава и регуляции численности популяций видов в сообществе.
Хищничество - форма межвидовых взаимоотношений, способ добывания пищи и питания животных (изредка - растений), при котором они (хищники) ловят, убивают и съедают других животных (жертв). В основе хищничества лежат пищевые связи. Хищничество встречается практически у всех животных, а также среди грибов (из гифомицетов) и у насекомоядных растений (росянка, пузырчатка и др.).
У хищников обычно хорошо развиты нервная система и органы чувств, позволяющие обнаружить и распознать свою добычу, а также средства овладения, поедания и переваривания добычи (острые втягивающиеся когти кошачьих, ядовитые железы многих паукообразных, стрекательные клетки актиний, длинные мягкие языки у жаб и ящериц и т.д.). По способу охоты хищников делят на засадчиков (подстерегающих жертву) и на преследователей. Иногда (например, у волков) встречаются коллективные формы охоты.
Не только у хищников есть специальные приспособления. У жертв тоже выработались защитные свойства в виде анатомо-морфологических, физиологических, биохимических и других особенностей. К ним можно отнести выросты тела, шипы, колючки, панцири, защитную окраску, ядовитые железы, способность быстро прятаться, быстро бегать, зарываться в грунт, строить недоступные хищникам убежища, прибегать к сигнализации об опасностях...
Между хищниками и жертвами существует равновесие. Для равновесия необходимы три звена. Например, олени поедают растения, а сами служат пищей волкам. Таким образом, плотность популяции растительноядного животного регулируется снизу (количество корма) и сверху (число хищников). При отсутствии волков дикие олени могут чрезмерно размножиться и подорвать свою кормовую базу, что затем вызовет гибель от голода значительной части их популяции. До недавнего времени существовало мнение, будто все хищники - вредные животные и их следует уничтожать. Но уничтожение хищников наносит большой ущерб природе. Ведь жертвами хищников обычно бывают больные и ослабленные особи, уничтожением которых сдерживается распространение болезней. Поэтому волки содействуют интенсивному размножению и повышению жизнеспособности популяций оленей. Таким образом, хищник - важнейший фактор естественного отбора.
Хищничество может являться причиной регулярных периодических колебаний численности популяций каждого из взаимодействующих видов. В природных условиях наблюдаются циклические колебания численности хищника и его жертвы; при этом циклы тесно связаны во времени: за подъемом численности жертв с некоторым запаздыванием следует подъем численности хищника, после которого численность жертв начинает падать. Классический пример таких циклов - колебания численности зайцев и рысей в Канаде. Было обнаружено, что численность зайца-беляка регулярно изменяется с периодом в 8-11 лет. Примерно с такой же периодичностью меняется численность рыси, причем максимум численности наступает на год-два позднее, чем у зайцев. Численность жертв лимитируется именно численностью хищников и наоборот (рис. 21).

Рис. 21. Отношения хищник-жертва поддерживают равновесие их популяций
Равновесие в системе хищник-жертва не возникает «автоматически», а устанавливается на протяжении многих сотен и даже тысяч лет. Но это равновесие может быть нарушено человеком при введении в систему слишком эффективного хищника, не свойственного данному биоценозу. Например, на острова человек завез домашних кошек, которые оказались значительно ловчее местных животных и в результате полностью истребили многие уникальные виды островной фауны. Завезенный хищник не обязательно сосредоточивается на истреблении своей привычной добычи. Например, лисы, интродуцированные в Австралию для уничтожения кроликов, нашли в изобилии более легкую добычу - местных сумчатых, не доставляя запланированной жертве особых хлопот.
Понятие «симбиоз» (от греч. symbiosis - сожительство) означает различные формы совместного существования организмов разных видов (симбионтов), с помощью которых эти организмы регулируют свои отношения с внешней средой. Выделяют следующие типы симбиоза: комменсализм, мутуализм, паразитизм, синойкия и другие.
Комменсализм, или нахлебничество, сотрапезничество (от лат. сот - совместно, сообща и mensa - стол), - форма симбиоза, при которой один из партнеров системы (комменсал) питается остатками пищи или продуктами выделения другого (хозяина), не причиняя последнему вреда. При комменсализме один из партнеров может использовать другого для защиты, как средство передвижения или питаться за его счет. Связи комменсалов могут быть условно разделены на несколько вариантов комменсализма.
«Нахлебничество» - потребление остатков пищи хозяина. Например, взаимоотношение львов с гиенами, подбирающими остатки недоеденной львами добычи. Краб пинникса (Pinnixa), поселяющийся в раковине дальневосточных пластинчатожаберных моллюсков (Pecten jessoensis), питается их объедками; мальки многих рыб живут среди щупальцев актиний и медуз и питаются отбросами их пищи. Песцы в тундре следуют за белым медведем и доедают остатки его пищи.
«Сотрапезничество» - потребление разных веществ или частей из одного и того же ресурса. Примером могут быть взаимоотношения почвенных бактерий-сапротрофов различных видов, перерабатывающих разные органические вещества перегнивших растительных остатков, и высших растений, потребляющих образовавшиеся при этом минеральные соли. Взаимосвязь клеста и белки в питании семенами хвойных растений также может служить примером «сотрапезничества».
Когда поедание пищи комменсалами начинает вредить хозяину, комменсализм переходит в конкуренцию или в паразитизм. Комменсализм тесно связан с синойкией.
Синойкия, или квартирантство (от греч. synoikia - совместное жилище), - форма симбиоза, разновидность комменсализма: совместное проживание двух организмов разных видов, полезное для одного и безразличное для другого.
В отличие от комменсализма при синойкии не возникает непосредственных пищевых отношений. Один из организмов может использовать другой как субстрат для заселения, средство перемещения и т.д. Такой тип взаимоотношений широко распространен у растений. Например, лианы и эпифиты, лишайники и мхи - на стволах и ветвях деревьев. В гнездах птиц и в норах грызунов обитает множество видов членистоногих. Некоторые виды насекомых обитают в муравейниках и термитниках.
Поверхностное размещение мелких организмов на более крупных называется эпойкией. Например, усоногие раки обитают на панцирях крабов, на коже китов и акул; рыбы-прилипалы - на теле акул; кишечнополостные - на раковинах моллюсков и т.д. Размещение мелких организмов внутри крупных называется энтойкия. Например, рыба фиерасфер (Fierasfer), живущая в водных легких голотурий (Holothuria tubulosa). Примером синойкии является форезия - перенос животными других, более мелких, слабоподвижных животных. Так, некоторые мучные клещи (Tyroglyphoidea) временно прикрепляются к телу более подвижных насекомых или мышей, крыс и с их помощью переселяются в новые места обитания. Взаимоотношения типа синойкии широко распространены у животных с растениями. Использование животными растительного субстрата для постройки жилищ (гнезда птиц, обитание в дуплах, в расщелинах коры), перенос семян и плодов растений животными - это примеры различных форм синойкии. Нередко синойкия становится начальным этапом более тесных взаимоотношений типа мутуализма и паразитизма.
Мутуализм (от лат. mutuus - взаимный) - высшая форма симбиоза, взаимовыгодное сожительство разных видов. При этом отношения между партнерами не только характеризуются взаимовыгодностью, но и ни один из них не может существовать без другого. Мутуализм делят на облигатный (обязательный для одного или для всех видов-партнеров) и факультативный (необязательный). Классическим примером мутуализма могут служить лишайники, которые представляют собой организмы, созданные симбиозом между водорослями и грибами.
Мутуалистические отношения высших растений с грибами представляет собой микориза (от греч. mykes - гриб и rhiza - корень) - грибокорень. Различают эктотрофную микоризу, при которой гриб оплетает корень, не проникая в клетки. Грибы пользуются выделениями корней и стимулируют усвояемость высшими растениями органических веществ почвы. При эндотрофной микоризе гифы гриба проникают в клетки корня растения и питаются как паразиты. При этом высшие растения получают дополнительное азотистое питание, а грибы - продукты фотосинтеза.
Взаимная польза от такого сожительства настолько велика, что многие высшие растения (ятрышники, вересковые, орхидеи) не могут расти без грибов, а грибы - без этих растений.
Важную роль играет и симбиоз растений с микробами, способными к азотофиксации. Например, в клетках корней некоторых растений (бобовые, березовые, лоховые, крушиновые и другие) поселяются почвенные бактерии, вызывая разрастание и формирование так называемых клубеньков. Клубеньковые бактерии, питаясь тканями растений, фиксируют атмосферный азот в виде соединений, которые могут усваиваться растением. Так, клевер и люцерна накапливают в клубеньках азота от 150 до 300 кг/га. Посевы бобовых применяются в сельском хозяйстве для обогащения почвы азотом.
Примером мутуализма являются взаимоотношения узко специализированных к опылению растений (инжир, дурман, купальница, орхидеи) с опыляющими их видами насекомых, летучих мышей и других животных. Приведем еще несколько примеров мутуализма. Кедровка, питающаяся только семенами кедровой сосны, является единственным распространителем ее семян. Птицы кормятся насекомыми-паразитами на колее носорога, а их взлет служит ему сигналом опасности. Термиты и живущие у них в кишечнике жгутиконосцы, обладающие способностью переваривать поглощаемую термитами клетчатку, которую эти насекомые без симбионтов не способны усваивать. Форму мутуализма приобретают отношения многих муравьев и тлей: муравьи защищают тлей от врагов, а сами питаются сахаристыми выделениями.
Классический пример мутуализма - сожительство рака-отшельника (Pagurus bernhardus) и актинии (Sagartia parasitica). Своеобразен мутуализм многих глубоководных рыб со светящимися бактериями. Например, у глубоководных удильщиков (Chaenophryne draco) ловчий орган имеет железу, в которой поселяются палочковидные бактерии, излучающие свет.
Мутуализм широко распространен в природе. Он сыграл огромную роль в возникновении и эволюции большинства современных форм жизни. В свете симбиотической теории образования эукариот (теории симбиогенеза) клетки практически всех эукариот - мутуалистические сообщества, состоящие из двух и трех компонентов (цитоплазмы, митохондрий и хлоропластов).
Паразитизм (от греч. parasites - нахлебник, дармоед) - межвидовые взаимоотношения (одна из форм симбиоза - совместного проживания), при которых один вид (паразит) использует другой (хозяина) как среду жизни и как источник пищи. Паразитизм возник в процессе тесного контакта разных видов организмов на основе пищевых и пространственных связей и встречается на всех уровнях организации живого, начиная с вирусов и бактерий и заканчивая высшими растениями и млекопитающими.
Таблица 5.1 Сводная таблица. Виды биотических взаимосвязей
| Тип связи | Вид связи | Определение | Примеры |
| 1 | 2 | 3 | 4 |
| Взаимополезные отношения | Симбиоз (++) | Взаимосвязи в виде сожительства с обоюдной пользой для симбионтов, но с некоторыми элементами паразитирования какого-то компонента (от греч. «симбиосис» - сожительство) | Сожительство гриба и водоросли (лишайники). Гриб и корни высших растений (микориза). Клубеньковые бактерии и корни бобовых растений |
| Мутуализм (++) | Взаимосвязь с обоюдной пользой, но без элементов паразитирования компонентов (от лат, «мутуо» - взаимно) | Энтомофилия. Эндозоохория (распространение семян плодоядными животными) | |
| Протокооперация (++) | Взаимосвязь, выгодная для обоих компонентов, но не обязательно должна быть (от греч. «протос» - первый и лат. «кооператно» - сотрудничество) | Вико-овсяные посевы. Совместная ловля рыбы бакланами и пеликанами | |
| Полезно-нейтральные отношения | Комменсализм (+0) | Взаимосвязь, при которой один компонент (хищник) получает какое-либо преимущество, не принося заметного вреда другому (от лат, «кум» - вместе и «менса» - стол, яства) | Взаимосвязь клеста и белки в питании семенами хвойных. Размещение эпифитов на ветвях деревьев. Акулы и рыбы прилипалы |
| Синойкия (+0) | Взаимосвязь, выгодная для одного, а для другого безразличная и необязательная(от лат. «син» - вместе и «ойкос» - жилище) | Экзохория. Постройка гнезд в кроне дерева | |
| Полезно-вредные отношения | Хищничество (+-) | Взаимосвязь, при которой один компонент поедает другого (жертву) | Волк - олень. Лиса - мыши. Поедание растений фитофагами. Поедание грибами (зоопагалами и монилиалами) нематод и амеб |
| Паразитизм (+-) | Взаимосвязь, при которой организмы одного вида живут за счет питательных веществ другого (хозяина), причем хозяин от этого не гибнет, а чувствует себя угнетенно | Наружный паразитизм, Внутренний паразитизм. Случайный паразитизм. Обязательный паразитизм | |
| Полупаразитизм (+-) | Взаимосвязь, при которой организм одного вида получает некоторые питательные вещества от хозяина, но и сам образует органические вещества | Иван-да-Марья и лесные травы. Омела и яблоня (или другие деревья) | |
| Аменсализм (+-) | Взаимосвязь, полезная для одного, но угнетающе действующая на другого (от лат. «а» - не, «менса» - стол, яства) | Угнетающее действие с помощью химических веществ (фитонциды, антибиотики) |
Паразитизм является одной из форм антагонистического симбиоза. Система «паразит-хозяин» построена по принципу неустойчивого равновесия, нарушение которого может привести к распаду системы и гибели одного или обоих партнеров.
Паразитов подразделяют на облигатных (обязательных) и факультативных (необязательных). Паразитизм может быть временным, когда паразиты нападают на хозяев только для питания (комары, слепни, вампиры и др.) и постоянным (стационарным), когда паразиты проводят на хозяине большую часть жизни (клещи, блохи, вши и другие). Иногда в процессе развития паразит меняет своих хозяев. Например, малярийный плазмодий часть жизни проводит в теле комара, а другую часть - в теле человека. Паразитов делят также на эктопаразитов, обитающих на поверхности тела хозяина, и эндопаразитов, живущих во внутренних полостях, тканях и клетках хозяина.
У паразитов выработались приспособления к паразитизму - упрощение организации, редукция органов. Например, у паразитических червей редуцированы органы пищеварения. У растений-паразитов сокращается количество хлорофиллоносной ткани, исчезают некоторые органы. Так, у повилики нет корней и зеленых листьев, она питается только за счет растения-хозяина. Некоторые организмы используют хозяина для откладывания яиц и выкармливания потомства. Например, самка насекомого афелинуса откладывает яйцо в тело тли, где развивающаяся личинка питается внутренним содержимым, уничтожая хозяина. Гнездовой паразитизм наблюдается у кукушки, подкладывающей свои яйца в гнезда другим птицам, перекладывая на них обязанности вскармливания птенцов.
Многие паразиты вызывают тяжелые заболевания у человека и животных, отравляя организм продуктами жизнедеятельности или истощая его. Изучение различных особенностей паразитизма важно для ведения борьбы с паразитами (табл. 5.1).
Лекция 13.
Экологическая ниша
Экологическая ниша - это положение вида, которое он занимает в общей системе биоценоза, комплекс его биоценотических связей и требований к абиотическим факторам среды. Понятие «экологическая ниша» ввел американский зоолог Дж. Гриннел для определения роли, которую играет тот или иной вид в биоценозе. Экологическая ниша - это абстрактное понятие, это совокупность всех факторов среды, в пределах которых возможно существование вида в природе. По образному выражению Ю. Одума, термин «экологическая ниша» относится к роли, которую играет организм в экосистеме; таким образом, местообитание - это «адрес», а ниша - «профессия» вида. Например, травоядные обитатели степей, антилопы и кенгуру, занимают одну экологическую нишу, но имеют различные места обитания. Наоборот, обитатели леса - белка и лось, относящиеся также к растительноядным животным, занимают разные экологические ниши.
Процесс разделения популяциями видов пространства и ресурсов называется дифференциацией экологических ниш. Наиболее ярко выражен процесс дифференциации экологических ниш у животных, что связано с разными видами рационов питания. Например, одни птицы питаются плодами и семенами растений, другие птицы ловят разных насекомых и червей. Хотя растения имеют один и тот же тип питания - автотрофный (фотосинтез) и используют одни и те же ресурсы среды (раствор минеральных веществ, диоксид углерода и солнечный свет), но и у них ниши дифференцированы. Есть растения светолюбивые и тенелюбивые; их корневые системы расположены на разной глубине; им требуется разное количество воды; они в разное время цветут и плодоносят; имеют разных опылителей. Ярусность в лесу - это пример разделения экологических ниш разных организмов (рис. 22).

Рис. 22. Экологические ниши некоторых птиц, населяющих хвойные леса
У организмов каждого вида может быть две ниши: фундаментальная и реализованная. Фундаментальная ниша - это те условия, в которых вид, в принципе, может существовать, а реализованная ниша - это те условия, где вид реально встречается в данном сообществе. Реализованная ниша всегда составляет некоторую часть фундаментальной.
Экологическая сукцессия
Сообщества формируются и изменяются в течение определенного времени. При этом изменяется их видовой состав, обилие разных групп организмов, трофическая структура, продуктивность и все остальные показатели. Последовательная смена одного биоценоза другим называется экологической сукцессией (от лат. succession - последовательность, смена). В целом цепь сменяющих друг друга биоценозов называется сукцессионным рядом или серией (сериальные стадии). В сукцессионном ряду каждый сериальный биоценоз представляет собой определенную стадию формирования конечного сообщества, называемого климаксным (от греч. climax - лестница) или климаксом. Примером сукцессии, приводящей к смене одного сообщества другим, может служить зарастание небольшого озера с последующим появлением на его месте болота, а затем - леса.
Учение о сукцессии разработали американские ботаники Г. Коулес и Ф. Клементс. Основная позиция Ф. Клементса заключается в том, что изменения во времени - естественное свойство сообществ. Первопричиной смены фитоценозов он считал изменение отдельных климатических факторов или их комплекса, а реакция экосистем в виде смены последовательного ряда сообществ представляет адаптивный ответ на экосистемном уровне. Сукцессия, по Ф. Клементсу, завершается формированием сообщества, наиболее адаптированного по отношению к комплексу климатических условий. Такое сообщество он назвал «климаксформация», или просто климакс.
Современная концепция экологических сукцессий расходится с утверждением Ф. Клементса об исключительном значении климата как движущей силы сукцессии. Смены сообществ могут происходить под влиянием других факторов, таких, как изменение рельефа, почвы, гидрологического режима и т.д. В современной экологии различают сукцессии экзогенетические и эндогенепгические. Экзогенетические сукцессии могут быть вызваны действием внешних, абиотических, факторов, или воздействием человека на биоценозы: мелиоративное осушение болот, загрязнение водоемов, неумеренный выпас скота и т.п. Эндогенетические сукцессии вызываются изменением структуры и системы связей в существующих сообществах. Впрочем, эти две категории сукцессий взаимосвязаны и могут переходить одна в другую, на что указывал академик В.Н. Сукачев.
По общему характеру сукцессии подразделяются на первичные и вторичные.
Первичная сукцессия начинается на месте, абсолютно лишенном жизни: вулканических туфовых и лавовых полях, сыпучих песках, скалах, каменистых россыпях. Здесь велика роль первых поселенцев: бактерий, лишайников, водорослей. Выделяя продукты своей жизнедеятельности, они изменяют материнскую породу, разрушая ее, и способствуют почвообразованию. Постепенное накопление отмерших, разлагающихся организмов и эрозия горной породы в результате выветривания формируют слой почвы для поселения мхов. Мхи по мере роста и развития продолжают процесс образования почвы. Формируется примитивное сообщество, способное существовать в неблагоприятных условиях, обеспечивающее все большее разнообразие организмов.
Вторичные сукцессии развиваются на месте, где ранее уже существовал хорошо развитый биоценоз. Например, на месте сгоревшего леса, заброшенного поля, осушенного болота или иного нарушенного сообщества. В таких местах обычно сохраняются богатые жизненные ресурсы. Например, на пожарище поселяются светолюбивые растения. Под их пологом начинают развиваться теневыносливые виды. Появление растительности улучшает состояние почвы, на которой поселяются новые виды, вытесняя первых поселенцев. В зависимости от почвы вторичная сукцессия может идти быстро или медленно, пока, наконец, не образуется климаксовое сообщество. Но скорость изменений при вторичной сукцессии гораздо выше, чем при первичной.
Пример продолжительности основных стадий вторичной сукцессии в умеренном климате:
- первая - стадия травянистой растительности - длится около 10 лет;
- вторая - стадия кустарников - длится от 10 до 25 лет;
- третья - стадия лиственных деревьев - длится от 25 до 100 лет;
- четвертая - стадия хвойных деревьев - длится более 100 лет.
Сукцессия - закономерный и направленный процесс, охватывающий общие изменения, происходящие на определенной стадии, свойственные любому сообществу. В качестве основных типов сукцессионных изменений выделяют следующие:
- виды растений и животных в процессе сукцессии непрерывно сменяются;
- сукцессионные изменения всегда сопровождаются повышением видового разнообразия организмов;
- происходит увеличение биомассы органического вещества;
- изменения состоят в снижении чистой продукции сообщества и повышении его дыхания, что является наиболее важным явлением сукцессии.
Лекция 14.
ЭКОЛОГИЯ ЭКОСИСТЕМ
Биогеоценоз и его структура
В природе популяции разных видов интегрируются в биоценозы. Но никакой биоценоз не может развиваться сам по себе, вне среды и независимо от нее. В результате в природе складываются определенные комплексы, совокупности живых и неживых компонентов. Пространство с более или менее однородными условиями, заселенные тем или иным сообществом организмов (биоценозом), называется биотопом. Биотоп - это место существования, местообитание биоценоза. Поэтому биоценоз можно рассматривать как исторически сложившийся комплекс организмов, характерный для какого-то конкретного биотопа. Любой биоценоз образует с биотопом биологическую систему более высокого ранга, чем биоценоз - макросистему - биогеоценоз. Биогеоценоз - исторически сложившаяся совокупность живых организмов (биоценоз) и абиотической среды вместе с занимаемым ими участком земной поверхности (биотопом) (рис. 23). Граница биогеоценоза устанавливается по границе растительного сообщества (фитоценоза) - важнейшего компонента биоценозов. Для каждого биогеоценоза характерен свой тип вещественно-энергетического обмена. Биогеоценоз - составная часть природного ландшафта и элементарная биотерриториальная единица биосферы.
Термин «биогеоценоз» предложил в 1944 году академик В.Н. Сукачев (1880-1967). По В.Н. Сукачеву, «биогеоценоз - это совокупность на известном протяжении земной поверхности однородных природных явлений (атмосферы, горной породы, растительности, животного мира и мира микроорганизмов, почвы и гидрологических условий), имеющая свою особую специфику взаимодействия этих слагающих ее компонентов и определенный тип обмена веществами и энергией между собой и другими явлениями природы и представляющая собой внутренне противоречивое единство, находящееся в постоянном движении, развитии». Формулируя понятие «биогеоценоз», В.Н. Сукачев объединил в нем структурную и функциональную значимость макросистемы.

Рис. 23. Схема структуры биогеоценоза
Биогеоценозы могут быть самых различных размеров: например, лес, озеро, луг и другие. Примером сравнительно простого биогеоценоза может служить небольшой водоем, пруд. Независимо от размеров и сложности любой биогеоценоз имеет следующий состав:
- продуценты - производители (зеленые растения),
- консументы - потребители (первичные - растительноядные животные, вторичные - плотоядные животные и т.д.),
- редуценты - разрушители (микроорганизмы),
- компоненты неживой природы.
Между ними возникают связи самых различных порядков. Устойчивость биогеоценозов поддерживается саморегуляцией.
Деятельность человека преобразовала природные биогеоценозы, и на их смену пришли агробиогеоценозы, количество которых постоянно увеличивается. Их примерами являются сельскохозяйственные поля, сады, пастбища, полезащитные лесные полосы, пруды и водохранилища, каналы, осушенные болота. Агробиогеоценозы по своей структуре характеризуются незначительным числом видов, но высокой их численностью. В настоящее время агроценозами занято более 10% суши Земли. Если природные биогеоценозы - саморегулируемые системы, то агроценозы регулируются человеком.
Экосистема и типы экосистем
В экологии наряду с термином «биогеоценоз» используется термин «экосистема». Экосистема (от греч. oikos - жилище, местообитание, объединение), экологическая система - это совокупность совместно обитающих организмов и условий их существования, находящихся в закономерной взаимосвязи друг с другом и образующих систему взаимообусловленных биотических и абиотических явлений и процессов. Экосистема - первичная единица биосферы.
Термин «экосистема» предложен английским ботаником А.Д. Тенсли (1871-1955). А. Тенсли считал, что экосистемы, «с точки зрения эколога, представляют собой основные природные единицы на поверхности Земли», в которые входит «не только комплекс организмов, но и весь комплекс физических факторов, образующих то, что мы называем средой биома, - факторы местообитания в самом широком смысле». Он подчеркивал, что для экосистем характерен «разного рода обмен веществ не только между организмами, но и между органическим и неорганическим».
Между «экосистемой» и «биогеоценозом» обычно ставят знак равенства, однако они несколько отличаются друг от друга. «Экосистема» - более широкое понятие, применяемое как к естественным разным по размерам комплексам (например, океан, тундра, лес, лужа, муравейник, капля воды и т.д.), так и к искусственным (например, аквариум, кабина космического корабля, территория фермы, город). Размерность экосистемы определяется содержанием исследуемого сообщества и его среды. Границы биогеоценоза чаще всего определяются закономерным сочетанием растительных сообществ однородного видового состава и строения. Биогеоценоз является одним из вариантов экосистемы. Однако существенных различий между экосистемой и биогеоценозом практически нет, и в последние годы наиболее употребительным становится термин «экосистема».
Между экосистемами, как и между биогеоценозами, обычно нет четких границ, и одна экосистема постепенно переходит в другую. Большие экосистемы состоят из экосистем меньшего размера, подобно «матрешкам», входя одна в другую. Например, муравейник, пень, нора с ее населением (микроэкосистема) входят в состав лесной экосистемы (мезоэкосистема). Лесная экосистема наряду с такими экосистемами, как луг, водоем, пашня, входит в состав более крупных экосистем - водосборный бассейн, природная зона, физико-географический район (макроэкосистема). Все экосистемы земного шара связаны через атмосферу и Мировой океан и составляют единое целое - биосферу - глобальную экосистему.
Экосистемы очень разнообразны. Их состав зависит от многих факторов, в первую очередь от климата, геологических условий и влияния человека. Если главную роль играют автотрофные организмы - продуценты (например, леса, луга, степи, болота и др.), системы называют автотрофными. Если же продуцентов в экосистеме нет либо их роль незначительна (например, экосистемы океанических глубин, высокогорных ледников), то такие экосистемы называют гетеротрофными. Различают наземные и водные экосистемы. Экосистемы могут быть как естественными, так и антропогенными (от греч. anthropos - человек и genos - происхождение), т.е. созданными человеком. Как, например, сельскохозяйственные, городские, промышленные экосистемы. Важнейшими природными экосистемами Земли являются: тайга; тундра и полярные пояса; океаны; побережья; болота, топи и мангровые заросли; степи умеренных широт; леса умеренных широт, саванны; пустыни; влажные экваториальные леса; горы; острова и другие.
Независимо от степени сложности экосистема характеризуется видовым составом, численностью входящих в нее организмов, биомассой, соотношением отдельных трофических групп, интенсивностью процессов продуцирования и деструкции органического вещества. Выделяют два подхода к изучению экологической системы: аналитический, при котором изучают отдельные части системы, и синтетический, когда изучают всю систему в целом.
Круговорот веществ и поток энергии в экосистемах
Вне зависимости от величины и степени сложности экосистемы являются открытыми системами и в большей или меньшей степени требуют постоянного притока энергии и различных веществ. В процессе жизнедеятельности организмов происходит постоянный приток энергии и круговорот веществ, причем каждый вид использует лишь часть содержащейся в органических веществах энергии. Происходит этот процесс через цепи питания (трофические уровни), представляющие собой последовательность видов, извлекающих органические вещества и энергию из исходного пищевого вещества; при этом каждое предыдущее звено становится пищей для следующего (рис. 24).
Круговорот веществ - это перемещение вещества в форме химических элементов и их соединений от продуцентов к редуцентам, через консументы или без них и опять к продуцентам. Растения - автотрофные организмы, способные в процессе фотосинтеза синтезировать органические вещества из неорганических, поэтому их называют продуцентами, или производителями.

Рис. 24. Поток энергии и круговорот веществ в экосистеме
Растения используются в качестве пищи животными, которые сами не способны к синтезу органики из неорганики. Такие гетеротрофные организмы называют консументами, или потребителями. Бактерии и грибы выполняют главную роль в разложении отмершей органики на исходные неорганические вещества, возвращая их в среду. Поэтому их называют деструкторами или редуцентами, т. е. разрушителями или восстановителями.
Итак, органическое вещество, образованное растениями, переходит в тело животных, а затем при участии бактерий вновь превращается в неорганические вещества, усваиваемые растениями. Таким образом в экосистеме осуществляется круговорот веществ.
Поток энергии - переход энергии в виде химических связей органических соединений (пищи) по цепям питания от одного трофического уровня к другому (более высокому) (рис. 25). Солнце является единственным источником энергии на Земле. Оно обеспечивает постоянный, непрерывный, незамкнутый приток энергии на Землю. В отличие от веществ, которые циркулируют по звеньям экосистемы и входят в круговорот, используясь многократно, энергия может быть использована только один раз.
Для понимания процессов потока энергии в экосистемах важно знать законы термодинамики. Первый закон термодинамики гласит, что энергия не может создаваться заново и не исчезает, а только переходит из одной формы в другую. Поэтому энергия в экосистеме не может появиться сама собой, а поступает в нее извне - от Солнца.

Рис. 25. Поток энергии в экосистеме
Второй закон термодинамики гласит, что процессы, связанные с превращениями энергии, могут протекать самопроизвольно лишь при условии, что энергия переходит из концентрированной формы в рассеянную. В соответствии с этим законом растениями используется лишь часть поступающей в экосистему солнечной энергии. Остальная энергия рассеивается и переходит в тепловую, которая расходуется на нагревание среды экосистемы. Небольшая часть солнечной энергии, поглощенная растением, расходуется на продукционный процесс, т. е. образование биомассы. Далее, переходя на следующие трофические уровни, вместе с пищей в виде химических связей, энергия также рассеивается и уменьшается в количестве, пока полностью не рассеется.
Пищевая цепь - основной канал переноса энергии в экосистеме. Растения являются первичными поставщиками энергии для всех других организмов в цепях питания. Существуют определенные закономерности перехода энергии с одного трофического уровня на другой вместе с потребляемой пищей. Во-первых, основная часть энергии, усвоенная консументом с пищей, расходуется на его жизнеобеспечение (движение, поддержание температуры и т.п.). Эту часть энергии рассматривают как траты на дыхание. Во-вторых, часть энергии переходит в тело организма потребителя «в запас». В-третьих, некоторая доля пищи не усваивается организмом, следовательно, из нее не высвобождается энергия. В последующем она высвобождается из экскрементов, но другими организмами (деструкторами), которые потребляют их в пищу. Выделение энергии с экскрементами у хищников невелико, у травоядных оно более значительно. Например, гусеницы некоторых насекомых, питающиеся растениями, выделяют с экскрементами до 70% энергии.
В каждом звене пищевой цепи большая часть энергии расходуется в виде тепла, теряется, что ограничивает число звеньев. В среднем, максимальные траты на дыхание в сумме с неусвоенной пищей составляют около 90% от потребленной. Поэтому переход энергии с одного трофического уровня на другой составляет всего около 10% энергии, употребленной в пищу. Нетрудно подсчитать, что энергия, доходящая до 5 уровня, составляет всего 0,01% энергии, поглощенной продуцентами. Эта закономерность называется «правилом десяти процентов». Она показывает, что цепь питания имеет ограниченное число звеньев, обычно не более 4-5. Пройдя через них, практически вся энергия оказывается рассеянной. Поэтому необходим постоянный приток энергии, чтобы экосистема могла существовать.
Лекция 15.
Биологическая продуктивность экосистемы
Биологическая продуктивность - это способность экосистемы к воспроизведению биомассы растений, микроорганизмов и животных, входящих в ее состав.
Биологическую продуктивность можно выразить продукцией - она является мерой биологической продуктивности и представляет собой количество воспроизведенной биомассы на 1 кв. м площади или в 1 куб. м объема в единицу времени и выражается в граммах углерода или сухого органического вещества.
Биомасса и продукция - основные показатели экосистемы. Биомасса, или урожай на корню, - масса органического вещества, заключенного в телах живых организмов на единицу площади. Биомасса может быть как живой, так и мертвой (например, древесина). Общая биомасса живого вещества составляет, по разным подсчетам, от 1800 до 2500 млрд. т (в среднем около 2000 млрд т) (таблица 6.1). Более 90% приходится на биомассу наземных растений (фитомассу), остальное - на водную растительность и гетеротрофные организмы. Таким образом, основная роль в живом веществе Земли принадлежит автотрофным растениям суши.
Основные запасы фитомассы приходятся на тропические области - более 55% (700-1000 т/га). На полярные области и пустыни приходится всего 1-2% (1-2 т/га). Биомасса животных (зоомасса) составляет около 5% всей биомассы. Биомасса Мирового океана в несколько сот раз меньше, чем биомасса суши. Причем здесь наблюдается обратное соотношение запасов биомассы растений и животных. Фитомасса океана составляет 0,2-0,3 млрд т, а зоомасса – 5-6 млрд т.
Важнейшим показателем экосистемы, кроме биомассы, является продукция. Продукция - прирост биомассы за единицу времени. Различают первичную и вторичную продукцию. Первичная продукция - органическое вещество, которое образуют продуценты в процессе фотосинтеза (или хемосинтеза). Вторичная продукция - биомасса, образуемая за единицу времени всеми репродуцентами и консументами. Общая годовая продукция наземной растительности оценивается приблизительно в 180-200 млрд т, основная ее доля приходится на тропическую зону. Годовая продукция океана составляет около 50-100 млрд т, т.е. он дает 1/3 всей продукции биосферы.
Дождевые тропические леса, характеризующиеся максимальной продуктивностью 20-25 т/га/год и биомассой 700-1000 т/га, рассматривают как основные аккумуляторы углерода и источники обогащения атмосферы кислородом и называют «легкими планеты».
Таблица 6.1 Биомасса организмов Земли (по Н.И. Базилевич и др.)
| Среда | Группа организмов | Масса 1012 т | Соотношение, в % |
| Континенты | Зеленые растения | 2,40 | 99,2 |
| Животные и микроорганизмы | 0,02 | 0,8 | |
| ИТОГО | 2,42 | 100,0 | |
| Океаны | Зеленые растения | 0,0002 | 6,3 |
| Животные и микроорганизмы | 0,0030 | 93,7 | |
| ИТОГО | 0,0032 | 100,0 | |
| Биомасса организмов Земли | 2,4232 |
Количественный учет потоков энергии и продуктивности экосистемы имеет большое практическое значение. Точный расчет потока энергии и продуктивности позволяет регулировать в экосистемах выход выгодной для человека биомассы живых организмов и хорошо представлять допустимые пределы ее изъятия.
Экологические пирамиды
Соотношение численностей, биомасс, и продукций (потоков энергии) популяций, относящихся к последовательным трофическим уровням, называют экологическими пирамидами. Их изображают в виде диаграмм, где ширина столбиков отражает соответствующий показатель каждого из уровней (рис. 26).

Рис. 26. Экологическая пирамида в общем виде
Эффект пирамиды в виде моделей придумал американский эколог Ч. Элтон. Экологические пирамиды бывают трех типов:
1. Пирамида численности отражает число организмов по трофическим уровням, причем численность особей при движении от продуцентов к консументам различного порядка значительно уменьшается. Например, на 1 га луга на I уровне находится около 9 млн. растений; на II уровне - 700 тыс. растительноядных насекомых; на III уровне - 350 тыс. хищных насекомых и пауков; на IV уровне - 3 птицы (рис. 27, а, б).

Рис. 27. Пирамиды численности
2. Пирамида биомасс показывает соотношение биомассы всех организмов в данной экосистеме по трофическим уровням (рис. 28). В наземных экосистемах масса продуцентов больше, чем масса консументов I порядка, консументов I порядка больше, чем консументов II порядка и т.д., поэтому график имеет вид нормальной пирамиды (рис. 29). В водных экосистемах экологическая пирамида перевернута (рис. 30), так как с повышением трофического уровня запас биомассы увеличивается.
Основной продуцент в водных экосистемах - фитопланктон. Одноклеточные водоросли, преобладающие в нем, живут от нескольких дней до нескольких недель. Организмы второго (потребитель планктона) и последующих трофических уровней живут дольше и накапливают большую биомассу. Такая перевернутая пирамида бывает не во все сезоны годы. Весной, в период массового развития планктона, его биомасса выше биомассы организмов второго и третьего трофических уровней.

3. Пирамида энергии - разновидность пирамиды биомасс, в которой представлено количество энергии, заключенной в каждом из трофических уровней экосистемы или проходящей через эти уровни (рис. 31).

Пирамиды потоков энергии никогда не бывают «перевернутыми», т.к. следующий трофический уровень может «пропустить через себя» лишь часть энергии, усвоенной предыдущим уровнем.
Лекция 16.
БИОСФЕРА
Учение В.И. Вернадского о биосфере
Впервые понятие «биосфера» (от греч. bios - жизнь и sphaira - шар) введено в биологию французским натуралистом Ж.Б. Ламарком в Париже в начале XIX столетия, а затем в геологию, уже в качестве термина - австрийским ученым Э. Зюссом (1875). Заслуга в разработке стройного, целостного научного учения о биосфере, как «области жизни», принадлежит академику В.И. Вернадскому (1863-1945). Под биосферой он понимал область существования живого вещества. Он назвал биосферой оболочку Земли, в формировании которой живые организмы играли и играют основную роль. «Жизнь захватывает значительную часть атомов, составляющих материю земной поверхности, - писал В.И. Вернадский. - Под ее влиянием эти атомы находятся в непрерывном, интенсивном движении. Из них все время создаются миллионы разнообразнейших соединений. И этот процесс длится без перерыва десятки миллионов лет, от древнейших археозойских эр до нашего времени. На земной поверхности нет химической силы, более постоянно действующей, а потому и более могущественной по своим конечным последствиям, чем живые организмы, взятые в целом».
Таким образом, живым организмам Вернадский отводил роль главнейшей преобразующей силы. При этом в понятие биосферы включается преобразующая деятельность организмов не только в границах распространения жизни в настоящее время (современная биосфера, или необиосфера), но и в прошлом (былые биосферы, или палеобиосферы). В таком случае под биосферой понимается все пространство (оболочка Земли), где существует либо когда-то существовала жизнь, т.е. где встречаются живые организмы или продукты их жизнедеятельности.
Современная биосфера представляет собой сложную систему, состоящую из многих компонентов, которые включают всю живую и неживую (среду обитания) природу. Она охватывает часть атмосферы, гидросферу и верхнюю часть литосферы, взаимосвязанные биогеохимическими циклами миграции веществ и энергии.
Границы биосферы. Верхняя граница биосферы в атмосфере определяется высотой 20-25 км - слоем озона; озоновый экран защищает все живое на Земле от ультрафиолетового излучения Солнца. Споры некоторых бактерий и плесневых грибов были обнаружены на высоте до 22 км.
В гидросфере границы доходят до максимальных глубин, жизнь встречается на дне океанических впадин, до 10-11 км от поверхности, где температура около 0°С.
В литосфере жизнь ограничивается прежде всего, температурой горных пород, постоянно возрастающей с глубиной. В породах земной коры бактерии были обнаружены на глубине 4 км.
В пределах биосферы выделяется биогеосфера («пленка жизни», «слой сгущения жизни», «биогеоценотический покров») - своеобразная оболочка земного шара, где сконцентрировано живое вещество планеты. В отличие от биосферы биогеосфера - это только область высокой концентрации живого вещества и располагается она на границе поверхности земной коры с атмосферой и в верхней части водной оболочки, занимая слой толщиной от нескольких метров (тундра, степь, пустыня) до сотен метров (леса, моря).
В основу учения Вернадского о биосфере положено представление о планетарной геохимической роли живого вещества в образовании биосферы как продукта длительного превращения веществ и энергии в ходе геологического развития Земли. В пределах биосферы везде встречается либо само живое вещество, либо следы его деятельности: газы атмосферы; природная вода; запасы нефти, угля, известняка, глины, сланцы, торф и т.д.
До трудов Вернадского в геологических явлениях и эволюции верхних слоев литосферы (земной коры) первенство отводилось физико-химическим процессам выветривания. В.И. Вернадский показал первостепенную преобразующую роль живых организмов и обусловливаемых ими механизмов разрушения горных пород, круговорота веществ, изменения водной и атмосферной оболочек Земли.
Структура биосферы. Функции и свойства живого вещества
В.И. Вернадский считал, что вещество биосферы состоит из семи глубоко разнородных частей, но геологически не случайных: живое вещество, биогенное, косное, биокосное, вещество в радиоактивном распаде, вещество рассеянных атомов и вещество космического происхождения. Наиболее важными являются первые четыре.
Живое вещество. Этот термин введен в литературу В.И. Вернадским. Под ним он понимал совокупность всех живых организмов, выраженную через массу, энергию и химический состав.
Биогенное вещество (уголь, нефть, известняки, торф, сланцы и др.) - органические и органоминеральные вещества, созданные живыми организмами на протяжении геологической истории Земли и являющиеся источником чрезвычайно мощной энергии.
Косное вещество представляет субстрат или среду обитания живых организмов, хотя образовано процессами, в которых живое вещество не принимало участия.
Биокосное вещество образовано в результате синтеза живого и неживого (косного) вещества (кора выветривания, почвы, илы, природные воды, осадочные породы и другие). Соотношение живого и косного в биокосном веществе варьирует. Например, почва в среднем состоит из 93% косных и 7% органических веществ.
Живое вещество - основа биосферы, хотя и составляет крайне незначительную ее часть (0,01% от массы всей биосферы). По Вернадскому, живое вещество - это совокупность существующих (или существовавших в определенный отрезок времени) живых организмов, являющихся мощным геологическим фактором. Живое вещество биосферы химически и геологически является чрезвычайно активным. Различают пять основных функций живого вещества на нашей планете.
1. Энергетическая функция заключается в осуществлении связи биосферно-планетарных явлений с космическим излучением, с солнечной радиацией. В основе этой функции лежит фотосинтез зеленых растений, в процессе которого происходит аккумуляция солнечной энергии и перераспределение ее между отдельными компонентами биосферы. За счет накопленной солнечной энергии протекают все жизненные явления на Земле.
2. Газовая функция обусловливает миграцию газов и их превращение, обеспечивает газовый состав атмосферы. Преобладающая масса газов на Земле имеет биогенное происхождение. В процессе функционирования живого вещества, создаются основные газы: азот, кислород, углекислый газ, сероводород, метан и другие. С газовой функцией связывают два переломных периода в развитии биосферы. Первый из них относится ко времени, когда содержание кислорода в атмосфере достигло примерно 1% от современного уровня (первая точка Пастера), что произошло около 1,2 млрд лет назад. Это обусловило появление первых аэробных организмов. С этого времени восстановительные процессы дополнились окислительными. Второй переломный период связывают со временем, когда концентрация кислорода достигла примерно 10% от современной (вторая точка Пастера), что создало условия для синтеза озона и образования озонового слоя в атмосфере. Это обусловило выход организмов из воды на сушу. До этого времени функцию защиты организмов от губительных ультрафиолетовых лучей выполняла вода.
3. Окислительно-восстановительная функция связана с интенсификацией под влиянием живого вещества процессов окисления (благодаря обогащению среды кислородом) и восстановления, - когда идет разложение органических веществ при дефиците кислорода, что сопровождается образованием и накоплением сероводорода.
4. Концентрационная функция проявляется в способности организмов концентрировать в своем теле рассеянные химические элементы, повышая их содержание по сравнению с окружающей средой на несколько порядков. Состав живого вещества существенно отличается от состава косного вещества планеты. В нем преобладают легкие атомы водорода, углерода, кислорода, натрия, магния, серы, хлора, калия, фосфора, кальция. Концентрация этих элементов в теле живых организмов в сотни и тысячи раз выше, чем во внешней среде. Этим объясняется неоднородность химического состава биосферы. Результат концентрационной деятельности - залежи торфа, углей, известняков, мела, рудные месторождения и другие.
5. Деструкционная функция обусловливает процессы, связанные с разложением органических веществ после смерти, вследствие чего происходит минерализация органического вещества, т.е. превращение живого вещества в косное. В результате образуются также биогенное и биокосное вещества биосферы. Основной механизм этой функции связан с круговоротом веществ. Наиболее существенную роль в этом отношении выполняют низшие формы жизни - грибы, бактерии (деструкторы, редуценты).
Биологический и геологический круговороты
Все вещества на планете находятся в процессе круговорота. Солнечная энергия вызывает на Земле два круговорота веществ: большой (геологический, биосферный) и малый (биологический).
Большой круговорот веществ в биосфере характеризуется двумя важными моментами: он осуществляется на протяжении всего геологического развития Земли и представляет собой современный планетарный процесс, принимающий ведущее участие в дальнейшем развитии биосферы.
Геологический круговорот связан с образованием и разрушением горных пород и последующим перемещением продуктов разрушения - обломочного материала и химических элементов. Значительную роль в этих процессах играли и продолжают играть термические свойства поверхности суши и воды: поглощение и отражение солнечных лучей, теплопроводность и теплоемкость. Неустойчивый гидротермический режим поверхности Земли вместе с планетарной системой циркуляции атмосферы обусловливал геологический круговорот веществ, который на начальном этапе развития Земли, наряду с эндогенными процессами, был связан с формированием континентов, океанов и современных геосфер. Со становлением биосферы в большой круговорот включились продукты жизнедеятельности организмов. Геологический круговорот поставляет живым организмам элементы питания и во многом определяет условия их существования.
Главные химические элементы литосферы: кислород, кремний, алюминий, железо, магний, натрий, калий и другие - участвуют в большом круговороте, проходя от глубинных частей верхней мантии до поверхности литосферы. Магматическая порода, возникшая при кристаллизации магмы, поступив на поверхность литосферы из глубин Земли, подвергается разложению, выветриванию в области биосферы. Продукты выветривания переходят в подвижное состояние, сносятся водами, ветром в пониженные места рельефа, попадают в реки, океан и образуют мощные толщи осадочных пород, которые со временем, погружаясь на глубину в областях с повышенной температурой и давлением, подвергаются метаморфозу, т. е. «переплавляются». При этой переплавке возникает новая метаморфическая порода, поступающая в верхние горизонты земной коры и вновь входящая в круговорот веществ (рис. 32).

Рис. 32. Геологический (большой) круговорот веществ
Наиболее интенсивному и быстрому круговороту подвергаются легкоподвижные вещества - газы и природные воды, составляющие атмосферу и гидросферу планеты. Значительно медленнее совершает круговорот материал литосферы. В целом каждый круговорот любого химического элемента является частью общего большого круговорота веществ на Земле, и все они тесно связаны между собой. Живое вещество биосферы в этом круговороте выполняет огромную работу по перераспределению химических элементов, беспрерывно циркулирующих в биосфере, переходя и.ч внешней среды в организмы и снова во внешнюю сроду.
Малый, или биологический, круговорот веществ – это циркуляция веществ между растениями, животными, грибами, микроорганизмами и почвой. Суть биологического круговорота заключается в протекании двух противоположных, но взаимосвязанных процессов - создания органических веществ и их разрушения. Начальный этап возникновения органических веществ обусловлен фотосинтезом зеленых растений, т. е. образованием живого вещества из углекислого газа, воды и простых минеральных соединений с использованием энергии Солнца. Растения (продуценты) извлекают из почвы в растворе молекулы серы, фосфора, кальция, калия, магния, марганца, кремния, алюминия, цинка, меди и других элементов. Растительноядные животные (консументы I порядка) поглощают соединения этих элементов уже в виде пищи растительного происхождения. Хищники (консументы II порядка) питаются растительноядными животными, потребляя пищу более сложного состава, включающую белки, жиры, аминокислоты и другие вещества. В процессе разрушения микроорганизмами (редуцентами) органических веществ отмерших растений и останков животных, в почву и водную среду поступают простые минеральные соединения, доступные для усвоения растениям, и начинается следующий виток биологического круговорота (рис. 33).
Рис. 33. Биологический круговорот веществ
Биогеохимические циклы
Обмен веществом и энергией, осуществляющийся между разными структурными частями биосферы и обусловленный жизнедеятельностью организмов, называется биогеохимическим циклом. Все биогеохимические циклы составляют современную динамическую основу существования жизни, они взаимосвязаны, и каждый из них играет свойственную ему роль в эволюции биосферы. Продолжительность циклов круговорота тех или иных веществ различна. Время, достаточное для полного оборота всего углекислого газа атмосферы через фотосинтез, составляет около 300 лет; кислорода атмосферы через фотосинтез – 2000-2500 лет; азота атмосферы через биологическую фиксацию, окисление электрическими разрядами - примерно 100 млн лет; воды через испарение - около 1 млн лет.
В большом и малом круговоротах участвуют множество химических элементов и их соединений. Но важнейшими из них являются круговороты биогенных элементов - кислорода, углерода, воды, азота, фосфора, серы. Большое значение имеют круговороты токсических элементов - ртути и свинца. Кроме того, из большого круговорота в биологический поступают многие вещества антропогенного происхождения (ДДТ, пестициды, радионуклиды и др.). Миграцию веществ в биохимических циклах можно рассмотреть на примере углерода (рис. 34).
Углерод содержится в основном в атмосфере в виде двуокиси углерода (CO2). В состав органического вещества он включается в процессе фотосинтеза растений. Затем основная масса его поступает в пищевые цепи и накапливается в телах животных в виде различных углеводородных соединений. Для обеспечения процессов жизнедеятельности значительная часть органических веществ растений и животных разлагается в процессе дыхания с выделением CO2 в атмосферу. Мертвое органическое вещество разлагается особой группой организмов (в основном микробами и грибами) до исходных минеральных веществ и углекислоты (СО2), которая также возвращается в атмосферу. Некоторая часть углерода включается в большой, или геологический, круговорот между сушей и океаном. В последнем она также включается в круговорот, начинающийся с фотоситтаирующих организмов фитопланктона. Большая доля органического вещества и содержащегося в нем углерода, по выражению В.И. Вернадского, «ускользает» из круговорота и уходит в геологию (в ископаемое состояние) в виде угля, торфа, нефти и других горючих соединений. Другая часть таким же образом концентрируется в донных карбонатных отложениях океана. Этот углерод, как и углерод горючих ископаемых, в настоящее время в значительной мере высвобождается человеком, использующим эти вещества в качестве энергетических, строительных и других ресурсов.

Рис. 34. Схема круговорота углерода
Возникновение и развитие ноосферы
Эволюция органического мира на Земле прошла несколько этапов. Первый связан с возникновением биологического круговорота веществ в биосфере. Второй сопровождался формированием многоклеточных организмов, что усложнило циклическую структуру биосферы. Эти два этапа называют биогенезом (от греч. bios - жизнь, genesis - происхождение). Третий этап связан с появлением человеческого общества, под влиянием которого в современных условиях происходит дальнейшая эволюция биосферы и превращение ее в сферу разума - ноосферу (от греч. noos - разум, sphaira - шар).
Ноосфера - новое состояние биосферы, когда разумная деятельность человека становится главным фактором, обусловливающим ее развитие.
Термин «ноосфера» был введен французскими учеными Э. Леруа и П.Т. де Шарденом в 1927 году. Они характеризовали ноосферу как особый, надбиосферный «мыслительный пласт», который «окутывает планету». В 30-40-е годы В.И. Вернадский углубил и развил учение о ноосфере. Он писал: «Ноосфера есть новое геологическое явление на нашей планете. В ней впервые человек становится крупной геологической силой. Он может и должен перестраивать своим трудом и мыслью область своей жизни, перестраивать коренным образом, по сравнению с тем, что было раньше».
В.И. Вернадский понимал под ноосферой новый этап в развитии биосферы и призывал к разумному регулированию отношений человека и природы. Необходимые предпосылки для создания ноосферы:
1. Человечество стало единым целым. Сегодня событие, происшедшее в захолустном уголке любого континента или океана, отражается и имеет большие или малые последствия в ряде других мест, на всей поверхности Земли.
2. Преобразование средств связи и обмена информацией, обеспечивающих сегодня мгновенную ее передачу.
3. Реальное равенство людей - как необходимое условие ноосферы.
4. Рост общего уровня жизни, как условие реального равенства людей, а также возможность влияния народных масс на ход государственных и общественных дел.
5. Развитие энергетики, открытие и использование новых видов энергии, необходимых для подъема уровня жизни.
6. Исключение войн из жизни общества.
Создание этих предпосылок становится возможным в результате взрыва научной мысли в XX веке. Это обстоятельство соответственно приведет к трансформации биосферы в ноосферу. Становление ноосферы теснейшим образом связано с овладением всеми формами движения материи и созданием новых живых организмов с помощью методов и средств биотехнологии и генной инженерии. Идеи Вернадского сегодня широко обсуждают в научных кругах. Они становятся методологической основой для многих современных концепций и теорий, направленных на решение экологических проблем и сохранение человечества.
Биосфера существовала до появления на Земле человека, может существовать и без него. Но человек без биосферы существовать не может. Осознание глобальной экологической опасности, грозящей гибелью человечеству, заставило мировое сообщество искать новые пути выхода из создавшегося положения и привело к пониманию необходимости разработки концепции устойчивого развития. Концепция была принята на конференции ООН по окружающей среде и развитию (Рио-де-Жанейро, 1992), где была провозглашена необходимость перехода мирового сообщества на путь устойчивого развития, т. е. развития общества на базе экологически целесообразного природопользования, обеспечивающего высокое качество жизни для людей на протяжении ряда поколений.
Модель устойчивого развития предполагает:
1. Снижение материало- и энергоемкости производства, максимальное сокращение отходов, снижение оборота токсичных веществ и расширение использования возобновляемых ресурсов, включая источники энергии.
2. Переход к ценообразованию, учитывающему экологические критерии (цену ущерба окружающей среде) и стимулирующему использование новых, экономически безопасных ресурсо- и энергосберегающих технологий в сочетании с системой налогов и штрафов.
3. Содействие устойчивому ведению сельского хозяйства и развитию сельских районов через повышение продуктивности сельскохозяйственных культур, улучшение питательных свойств растительной и животной продукции, использование комплексных методов борьбы с вредителями сельского хозяйства и т.д.
4. Передачу индустриально развитыми странами передовых технологий развивающимся странам, в частности, новых технологий, созданных на основе генетических материалов, полученных из развивающихся стран.
5. Создание международных институтов, способных определить единую глобальную линию устойчивого развития, устанавливать единые для всех стран экологические стандарты, аккумулировать и перераспределять ресурсы в интересах всего сообщества, контролировать соблюдение всеми государствами единых правил экологического поведения.
Дата добавления: 2021-04-24; просмотров: 316; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!